
This Key Event Relationship is licensed under the Creative Commons BY-SA license. This license allows reusers to distribute, remix, adapt, and build upon the material in any medium or format, so long as attribution is given to the creator. The license allows for commercial use. If you remix, adapt, or build upon the material, you must license the modified material under identical terms.
Relationship: 3465
Title
Decreased, GnRH pulsatility/release leads to Decreased, LH Surge
Upstream event
Downstream event
Key Event Relationship Overview
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding | Point of Contact | Author Status | OECD Status |
|---|---|---|---|---|---|---|
| Decreased, GnRH pulsatility/release leading to estradiol availability, increased via impaired ovulation | adjacent | High | High | Martina Panzarea (send email) | Under development: Not open for comment. Do not cite |
Taxonomic Applicability
| Term | Scientific Term | Evidence | Link |
|---|---|---|---|
| mammals | mammals | NCBI |
Sex Applicability
| Sex | Evidence |
|---|---|
| Unspecific |
Life Stage Applicability
| Term | Evidence |
|---|---|
| All life stages |
Key Event Relationship Description
Evidence Collection Strategy
The development of the KER is based on structured literature review of records. Description for KER is based on reviews and books on the topic. The method used are described in Annex B.1.
Evidence Supporting this KER
Biological Plausibility
The major role of the GnRH is to interact with its receptor expressed by gonadotropic cells in the anterior pituitary inducing the release of the gonadotropins, luteinizing hormone (LH) and follicle-stimulating hormone (FSH). The biological plausibility to the key event relationship is supported by the physiological role of GnRH surge in evoking LH surge.
Empirical Evidence
In vivo
Atrazine
The effect of atrazine (showing aromatase inducing properties) in lowering LH surge was demonstrated in several papers (Foradori 2009, 2011, 2013, 2014). In Foradori et al., 2009, ovariectomized adult Wistar rats were administered atrazine (50, 100, or 200 mg/kg of body weight daily by gavage) or vehicle for 4 days. Animals were primed with estrogen and progesterone to induce an evening LH surge. Blood samples were obtained over the afternoon and evening and plasma was assayed for LH and FSH. Another cohort of animals was transcardially perfused in the afternoon to examine GnRH activation using FOS immunoreactivity. 4-day treatment with atrazine resulted in a significant reduction in the magnitude of the LH and FSH surges, and this corresponds to a decrease in GnRH neurons expressing FOS immunoreactivity. Examining LH levels and GnRH activation 2 days and 4 days after atrazine withdrawal, within 4 days (but not 2 days) after cessation of atrazine treatment, measures of hypothalamic-pituitary-gonadal (HPG) activation returned to normal (Fig. 4 and Fig. 5).
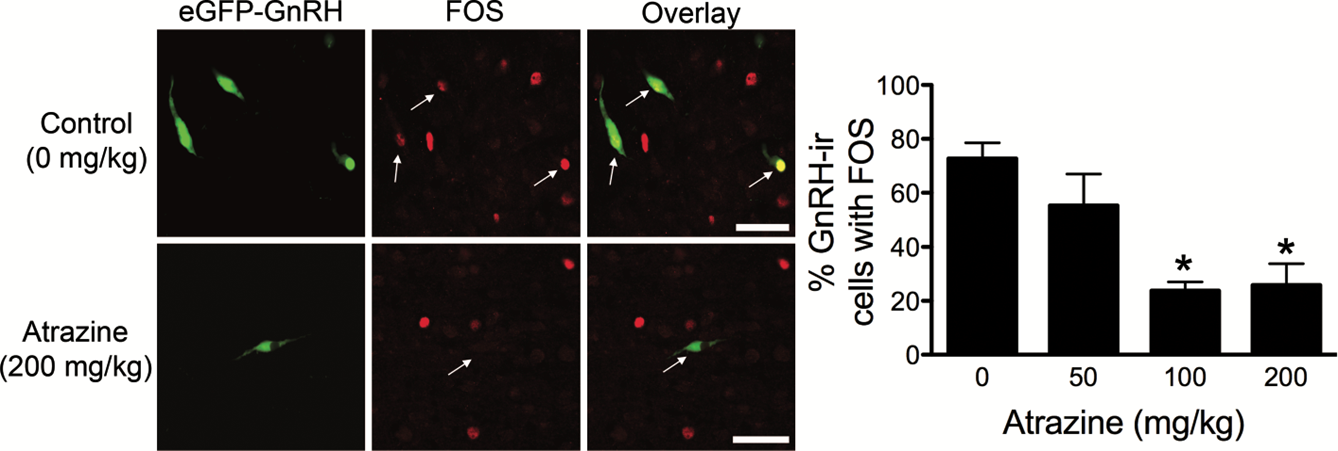
Figure 4. Photomicrographs of eGFP-GnRH (green), FOS (red), and overlay of control animals and animals treated with 200 mg/kg of atrazine; bar 1⁄4 50 lm. Histogram of the mean ± SEM percentage of GnRH neurons immunoreactive for FOS. *Significant difference from control and 50 mg/kg of atrazine (P < 0.05). GnRH-ir, GnRH immunoreactive. From Foradori et al., 2009
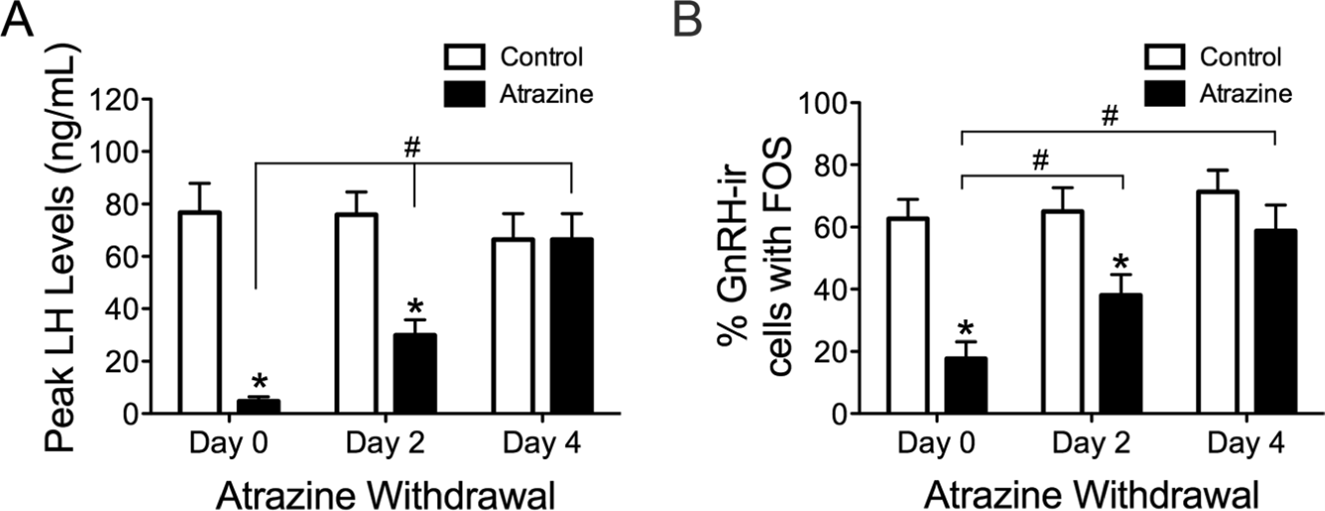
Figure 5. Histograms depicting the mean ± SEM peak LH levels (A) and the percentage of GnRH cells immunoreactive for FOS (B) on the final day of atrazine treatment (Day 0) and 2 days (Day 2) and 4 days (Day 4) after final atrazine treatment. *Significant difference from control group (p<0.05). #Significant difference between atrazine-treated groups (p<0.05). GnRH-ir, GnRH immunoreactive. From Foradori et al., 2009.
In Foradori et al., (2013) ATR treatment (200 mg/kg or vehicle daily for 4 days via gavage to ovariectomized adult female Wistar rats were) resulted in no changes to GnRH gene expression, peptide levels, or immunoreactivity but a reduction in GnRH pulse frequency and an increased pulse amplitude. These findings suggest that ATR acts to inhibit the secretory dynamics of GnRH pulses without interfering with GnRH mRNA and protein synthesis.
Atrazine failed also to alter pituitary sensitivity to the GnRH at any dose used.
Tributyltin (TBT)
Ovariectomized Wistar rats exposed to 100 ng TBT/kg bw/d for 15 days had reduced hypothalamic GnRH mRNA expression and exogenous KISS responsiveness, decreased basal and surge LH levels, reduced exogenous GnRH responsiveness, as well as decreased expression of both estrogen receptor (ER) (ERα and ERβ) in the pituitary (Fig. 6, Sena et al., 2017).
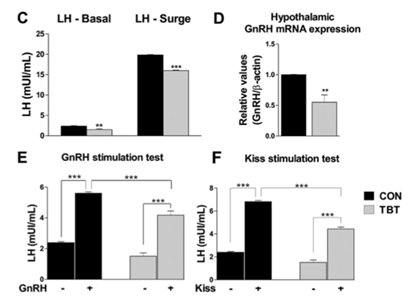
Figure 6. Tributyltin (TBT) rats have an abnormal estrous cycle. Proestrus (P), estrus (E), metestrus-diestrus (M-D). (C) Baseline and surge serum LH levels (n = 5–6). (D) qPCR analysis of GnRH mRNA extracted from female rat hypothalamic tissue (n = 4). (E) GnRH stimulation test. Assessment of serum LH levels 10 min after injection of GnRH agonist (0.1 nmol/kg via ip). Increased serum LH levels in rats treated with a GnRH agonist (indicated as GnRH +) were identified in both control and TBT rats (n = 5–6). (F) Kisspeptin stimulation test. Assessment of serum LH levels 10 min after injection of kisspeptin-10 (10 nmol ip). Increased serum LH levels in rats treated with kisspeptin (indicated as Kiss +) were identified in both control and TBT rats (n = 5–6). Significant differences compared **p ≤ 0.01 and ***p ≤ 0.01 vs saline-treated control groups (indicated as GnRH − or Kiss −). Two-way ANOVA indicated an interaction between groups and treatment (Tukey's multiple comparison test). From Sena et al., 2017.
As reported by Sena et al., (2017), TBT leads to a decreased LH and FSH secretion as result of a lower stimulatory effect induced by reduced GnRH expression levels.
A reduction in the hypothalamic GnRH mRNA expression, the impairment of Kisspeptin-stimulated GnRH release was also observed. As a consequence:
- a reduction in the serum E2 levels and an increase in the serum T levels, leading to abnormal regulation of feedback mechanisms.
- increased leptin secretion may also associate with the impairment of Kisspeptin-stimulated GnRH release.
Endopeptidases
The endopeptidase EC 3.4.24.15 (EP24.15) is a zinc metalloendopeptidase that is widely distributed in a variety of tissues, including the testes, pituitary and the central nervous system.
In recent studies peptidases acting on GnRH at the level of the median eminence have been isolated, the most important appearing to be the zinc metalloendopeptidase-24.15 (EP 24.15; 119 –121).
These observations suggest that endopeptidases exert a physiological role in shaping the GnRH signal directed at the pituitary gonadotrophs at the time of the LH surge and that this may be regulated by gonadal steroids including estrogen.
Ladsun et al., (1989) concluded that EP24.15 is the dominant factor determining the in vivo LHRH (former name of GnRH) degradation, suggesting a role of the enzyme in the regulation of the amount LHRH reaching pituitary gonadotropes (see also Molineaux et al., 1988).
EP 24.15 cleaves the central Tyr5-G1y6 bond in GnRH (Orlowski et al., 1983; Chu and Orlowski, 1985), and it is the primary responsible of the degradation of LHRH in hypothalamic and pituitary membrane preparations and in intact AtT20 cells (an anterior pituitary tumor cell line) (Molineaux, et at., 1988, see Table 1). After intracerebroventricular (icv). administration of GnRH, only about 1% of this peptide was recovered from brain after 1 hr. Concurrent administration of GnRH and N- [1-(RS)-carboxy-3-phenylpropyl]-Ala-Ala-Phe-p-aminobenzoate (cFP-AAF-pAB, a specific inhibitor of EP 24.15), led to a more than 10-fold increase in GnRH recovery. Administration of N-[1- (RS)-carboxy-3-phenylpropyl]-Phe-pAB(cFP-F-pAB) or captopril, inhibitors of other endopeptidases as “enkephalinase” (EP 24.1 1) and angiotensin converting enzyme respectively, did not significantly increase LH recovery.
Intravenous administration of GnRH and either cFP-F-pAB or cFP-AAF-pAB but not captopril, led to an increase in the half-life of GnRH from 10 min to 15 and 20 min, respectively. Concurrent administration of both inhibitors resulted in a dramatic 8-fold increase in the half-life of GnRH Concentrations of plasma GnRH 65 to 80 min after administration of inhibitors were 100- to 200-fold than those in controls.
Table 1. Inhibition of LHRH (GnRH) breakdown by peptidase inhibitors in rats. From Molineaux, 1988.

Wu et al., in 1997 (Fig. 7) demonstrated that Endopeptidase EC 3.4.24.15 in the Rat Median Eminence and Hypophysial Portal Blood modulates the LH surge (an inhibition of its activity with a specific inhibitor augmented the steroid-induced LH increase in ovariectomized rats).
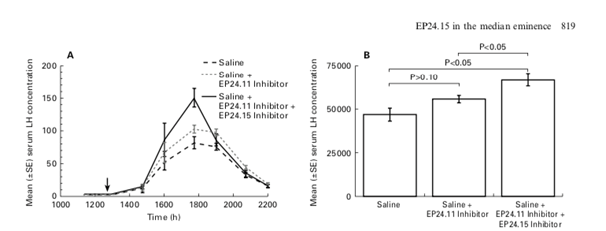
Figure 7. shows the mean (±SE) serum LH concentration (ng/ml) in rats treated with saline only (dashed line), saline plus the EP24.11 inhibitor (dotted line), or saline plus the EP24.11 inhibitor plus the EP24.15 inhibitor (solid line). (B) shows the mean (±SE) integrated area under the curve of serum LH concentration in rats administered saline alone, saline+EP24.11 inhibitor, and saline+EP24.11 inhibitor+EP24.15 inhibitor (from Wu et al., 1997).
GABA modulators
Muscimol (a selective agonist of GABAAR) significantly decreased GnRH biosynthesis in in the preoptic area (POA, anterior (AH) and ventromedial (VMH) hypothalamus, stalk/median eminence (SME), and led to analogous changes in plasma LH concentration (Ciechanowska et al., 2019, see Fig. 8 and 9).
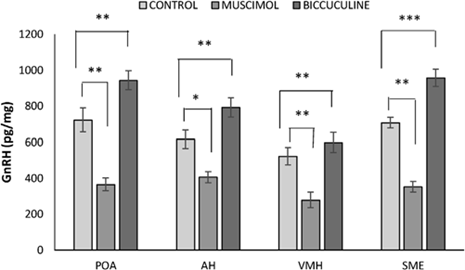
Figure 8. Effect of muscimol and bicuculline on the level of GnRH in the preoptic area (POA), anterior hypothalamus (AH), ventromedial hypothalamus (VM) and the stalk median eminence (SME) of follicular-phase ewes, *p < 0.05, **p < 0.01, ***p < 0.001. Asterisks indicate values that differ significantly from the control group animals. Data are the mean ± SEM, n=6 animals per group (one-way ANOVA) (from Ciechanowska et al., 2019).
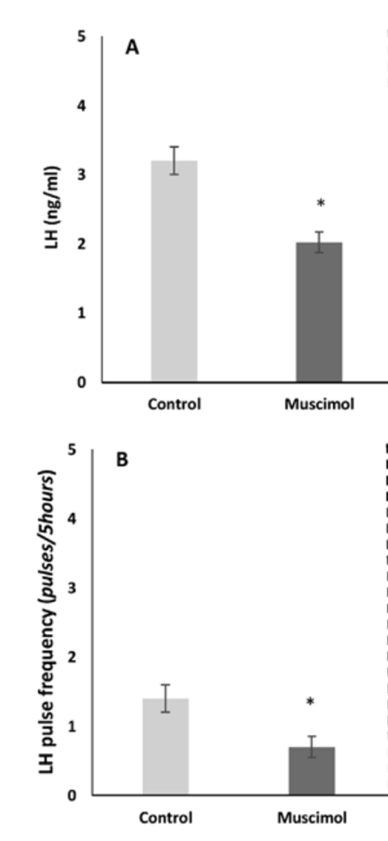
Figure 9. Effect of muscimol on the level of LH (A) and LH pulse frequency (B) in blood plasma of follicular-phase ewes, *p<0.05. Asterisks indicate values that differ significantly from the control group animals. Data are the mean SEM, n =6 animals per group (one way ANOVA, Wilcoxon test) (from Ciechanowska et al., 2019).
Light stimulation
In a proposed frequency-dependent control of GnRH neuron firing by RP3V neurons (Piet et al., 2018), neurons using GABA alone (green), kisspeptin alone (blue) and both transmitters innervate GnRH neuron cell bodies and proximal dendrites. At low firing frequencies (2 Hz), RP3V neuronal populations only release GABA that generates small transient increases in GnRH neuron firing (green) and has no substantial impact upon LH secretion (red). At higher firing frequencies (10 Hz), proposed to occur on the afternoon of proestrus, RP3V inputs to GnRH neurons now release kisspeptin (blue) to generate the preovulatory LH surge (red) (see also Fig. 10).
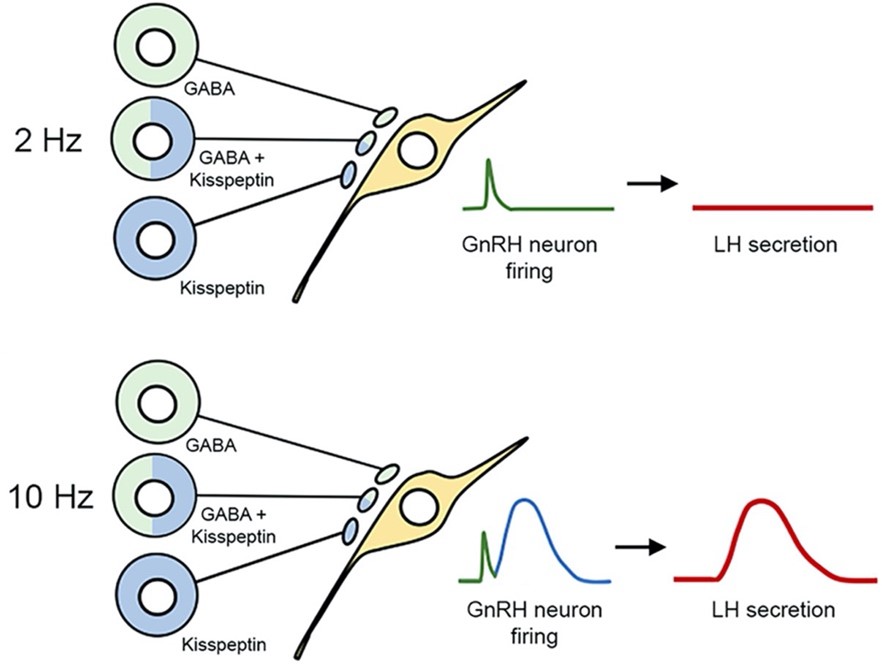
Figure 10. Schematic diagram showing proposed frequency-dependent control of GnRH neuron firing by RP3V neurons. Neurons using GABA alone (green), kisspeptin alone (blue) and both transmitters innervate GnRH neuron cell bodies and proximal dendrites (from Piet et al., 2018).
Dose concordance and temporal concordance
See Annex B.3.
Uncertainties and Inconsistencies
Atrazine
- the effective doses are about 150 times the NOAEL of atrazine (WHO, 2010)
Muscimol, TBT, Atrazine, endopeptidase
The Risk of Bias (RoB) of the primary research study was not evaluated.
For muscimol opposite effect reported when tested in vitro or in other in vivo models.
Known modulating factors
Not applicable/not investigated in detail.
Quantitative Understanding of the Linkage
At present we are unaware of any well-established quantitative relationships between Kea and KEb.
Response-response Relationship
Time-scale
Known Feedforward/Feedback loops influencing this KER
Domain of Applicability
References
Chu TG and Orlowski M, 1985. Soluble metalloendopeptidase from rat brain: action on enkephalin-containing peptides and other bioactive peptides. Endocrinology, 116:1418-1425. doi: 10.1210/endo-116-4-1418
Ciechanowska MO, Łapot M, Kowalczyk M, Malewski T, Brytan M, Antkowiak B and Przekop F, 2019. Does kisspeptin participate in GABA-mediated modulation of GnRH and GnRH receptor biosynthesis in the hypothalamic-pituitary unit of follicular-phase ewes? Pharmacol Rep, 71:636-643. doi: 10.1016/j.pharep.2019.02.019
Foradori CD, Hinds LR, Hanneman WH and Handa RJ, 2009. Effects of atrazine and its withdrawal on gonadotropin-releasing hormone neuroendocrine function in the adult female Wistar rat. Biol Reprod, 81:1099-1105. doi: 10.1095/biolreprod.109.077453
Foradori CD, Hinds LR, Quihuis AM, Lacagnina AF, Breckenridge CB and Handa RJ, 2011. The differential effect of atrazine on luteinizing hormone release in adrenalectomized adult female Wistar rats. Biol Reprod, 85:684-689. doi: 10.1095/biolreprod.111.092452
Foradori CD, Sawhney Coder P, Tisdel M, Yi KD, Simpkins JW, Handa RJ and Breckenridge CB, 2014. The effect of atrazine administered by gavage or in diet on the LH surge and reproductive performance in intact female Sprague-Dawley and Long Evans rats. Birth Defects Res B Dev Reprod Toxicol, 101:262-275. doi: 10.1002/bdrb.21109
Foradori CD, Zimmerman AD, Hinds LR, Zuloaga KL, Breckenridge CB and Handa RJ, 2013. Atrazine inhibits pulsatile gonadotropin-releasing hormone (GnRH) release without altering GnRH messenger RNA or protein levels in the female rat. Biol Reprod, 88:9. doi: 10.1095/biolreprod.112.102277
Ladsun A, Reznik S, Molineaux CJ and Orlowski M, 1989. Inhibition of endopeptidase 24.15 slows the in vivo degradation of luteinizing hormone-releasing hormone. J Pharmacol Exp Ther, 251:439-447
Molineaux CJ, Lasdun A, Michaud C and Orlowski M, 1988. Endopeptidase-24.15 is the primary enzyme that degrades luteinizing hormone releasing hormone both in vitro and in vivo. J Neurochem, 51:624-633. doi: 10.1111/j.1471-4159.1988.tb01084.x
Orlowski M, Michaud C and Chu TG, 1983. A soluble metalloendopeptidase from rat brain. Purification of the enzyme and determination of specificity with synthetic and natural peptides. Eur J Biochem, 135:81-88. doi: 10.1111/j.1432-1033.1983.tb07620.x
Piet R, Kalil B, McLennan T, Porteous R, Czieselsky K and Herbison AE, 2018. Dominant Neuropeptide Cotransmission in Kisspeptin-GABA Regulation of GnRH Neuron Firing Driving Ovulation. The Journal of neuroscience : the official journal of the Society for Neuroscience, 38:6310-6322. doi: 10.1523/jneurosci.0658-18.2018
Sena GC, Freitas-Lima LC, Merlo E, Podratz PL, de Araújo JFP, Brandão PAA, Carneiro MTWD, Zicker MC, Ferreira AVM, Takiya CM, de Lemos Barbosa CM, Morales MM, Santos-Silva AP, Miranda-Alves L, Silva IV and Graceli JB, 2017. Environmental obesogen tributyltin chloride leads to abnormal hypothalamic-pituitary-gonadal axis function by disruption in kisspeptin/leptin signaling in female rats. Toxicology and Applied Pharmacology, 319:22-38. doi: https://doi.org/10.1016/j.taap.2017.01.021
World Health Organization. Pesticide Residues in Food 2007: Toxicological Evaluations. Vol. 23. World Health Organization, 2010.
Wu TJ, Pierotti AR, Jakubowski M, Sheward WJ, Glucksman MJ, Smith AI, King JC, Fink G and Roberts JL, 1997. Endopeptidase EC 3.4.24.15 presence in the rat median eminence and hypophysial portal blood and its modulation of the luteinizing hormone surge. J Neuroendocrinol, 9:813-822. doi: 10.1046/j.1365-2826.1997.00637.x