
This AOP is licensed under the BY-SA license. This license allows reusers to distribute, remix, adapt, and build upon the material in any medium or format, so long as attribution is given to the creator. The license allows for commercial use. If you remix, adapt, or build upon the material, you must license the modified material under identical terms.
AOP: 342
Title
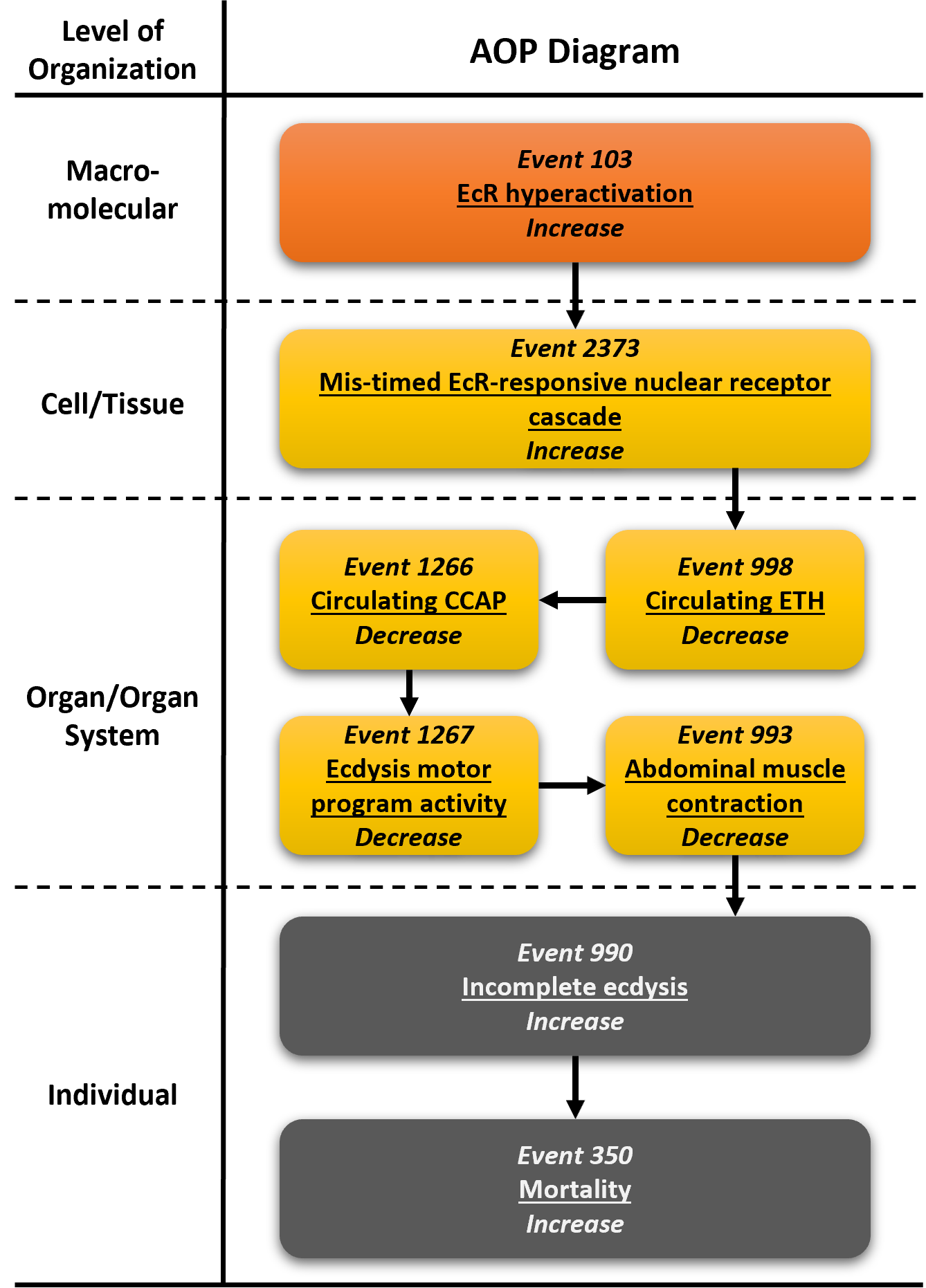
Ecdysone receptor hyperactivation leading to mortality via mis-timed activation of nuclear receptor cascade
Short name
Graphical Representation
Additional AOP Exploration Options
Click links below to explore AOP 342, Ecdysone receptor hyperactivation leading to mortality via mis-timed activation of nuclear receptor cascade in tools offered by third parties.
Point of Contact
Contributors
- You Song
Coaches
OECD Information Table
| OECD Project # | OECD Status | Reviewer's Reports | Journal-format Article | OECD iLibrary Published Version |
|---|---|---|---|---|
This AOP was last modified on October 01, 2025 16:00
Revision dates for related pages
| Page | Revision Date/Time |
|---|---|
| Increase, Ecdysone receptor hyperactivation | September 29, 2025 04:47 |
| Increase, Mis-timed ecdysone receptor-responsive nuclear receptor cascade | September 29, 2025 05:13 |
| Decrease, Circulating ecdysis triggering hormone | May 24, 2018 16:34 |
| Decrease, Circulating crustacean cardioactive peptide | May 24, 2018 16:37 |
| Decrease, Ecdysis motor program activity | September 29, 2025 04:18 |
| Decrease, Abdominal muscle contraction | May 24, 2018 16:41 |
| Increase, Incomplete ecdysis | May 24, 2018 16:41 |
| Increase, Mortality | October 26, 2020 05:18 |
| Increase, EcR hyperactivation leads to Increase, Mis-timed EcR-responsive NR cascade | September 29, 2025 04:55 |
| Increase, Mis-timed EcR-responsive NR cascade leads to Decrease, Circulating ETH | September 29, 2025 04:55 |
| Decrease, Circulating ETH leads to Decrease, Circulating CCAP | February 09, 2017 03:34 |
| Decrease, Circulating CCAP leads to Decrease, Ecdysis motor program activity | February 09, 2017 03:35 |
| Decrease, Ecdysis motor program activity leads to Decrease, Abdominal muscle contraction | September 29, 2025 04:17 |
| Decrease, Abdominal muscle contraction leads to Increase, Incomplete ecdysis | December 03, 2016 16:38 |
| Increase, Incomplete ecdysis leads to Increase, Mortality | December 03, 2016 16:38 |
| 20-hydroxyecdysone | February 09, 2017 03:06 |
| Tebufenozide | February 09, 2017 03:06 |
| Ponasterone A | February 09, 2017 03:06 |
| Methoxyfenozide | February 09, 2017 03:42 |
| Halofenozide | February 06, 2017 12:28 |
| Chromafenozide | February 09, 2017 03:41 |
| RH-5849 | February 09, 2017 03:43 |
| Muristerone A | October 01, 2025 07:04 |
Abstract
This AOP describes how hyperactivation of the ecdysone receptor (EcR) initiates a cascade of mis-timed nuclear receptor signaling, disrupting the tightly regulated endocrine control of insect molting. Dysregulation of key neuropeptides (ETH, CCAP) and subsequent impairment of motor programs and abdominal muscle contractions lead to incomplete ecdysis. Failure to properly shed the cuticle results in increased mortality.
This AOP highlights a critical endocrine mechanism relevant to insect growth and survival, providing a biologically plausible framework for linking molecular-level perturbations to population-relevant outcomes. It has direct regulatory relevance for evaluating endocrine-disrupting chemicals that target EcR signaling, such as insect growth regulators (IGRs), which are widely used in pest control but may pose risks to non-target insect populations, including pollinators and beneficial species.
By capturing key mechanistic events from receptor hyperactivation through behavioral and physiological impairment to organismal death, this AOP can support ecological risk assessment, guide the development of mechanistic testing strategies, and inform regulatory decision-making for chemicals acting on the EcR pathway.
AOP Development Strategy
Context
The development of this AOP was motivated by the regulatory and ecological importance of chemicals that target the ecdysone receptor (EcR) in insects. EcR is the molecular target of several classes of insect growth regulators (IGRs), which are deliberately designed to disrupt molting and cause lethality in pest species. However, these same mechanisms raise concern for potential adverse effects on non-target insects, including pollinators and beneficial arthropods. Because ecdysis is a highly conserved and hormonally regulated process essential for insect survival, disruptions of EcR signaling provide a biologically plausible and ecologically relevant pathway to mortality.
This AOP was therefore prioritized to support risk assessment and decision-making for EcR-active substances. It provides a mechanistic framework that integrates a large body of invertebrate endocrinology and neuroethology, allowing regulators to trace the cascade from molecular receptor activation to survival outcomes. The background understanding of ecdysis neuroendocrinology—developed over decades of model insect research—offers strong biological plausibility and cross-species relevance, making it a valuable case study for mechanistic regulatory science.
Strategy
The identification and evaluation of the key events (KEs) and key event relationships (KERs) were based on a structured strategy combining expert input, preliminary scoping, and systematic literature surveys:
-
Preliminary scoping and expert input
-
The starting point was established entomological literature on EcR signaling, molting neuroendocrine cascades, and motor program control.
-
Expert consultations with insect endocrinologists and toxicologists helped define the initial scope: perturbations to EcR signaling leading to lethal molting failure.
-
The AOP framework was cross-checked against existing knowledge of other insect hormone pathways (juvenile hormone, PTTH) to ensure specificity and avoid redundancy.
-
-
Literature identification and screening strategy
-
Databases searched: PubMed, Web of Science
-
Search terms: “ecdysone receptor agonist,” “EcR hyperactivation,” “ecdysis neuroendocrine cascade,” “ETH CCAP insect molting,” “ecdysis motor program,” “ecdysis mortality,” “insect growth regulator EcR,” “ecdysteroid signaling timing,” and combinations thereof.
-
Timeframe: Literature spanning ~1970 to 2023 was considered, as foundational insect endocrinology was established in the late 20th century and more recent studies provide mechanistic and toxicological data.
-
Searches emphasized both mechanistic biology (basic insect physiology and endocrinology) and applied ecotoxicology (chemical testing, IGR exposure studies).
-
-
Screening and selection
-
Abstracts were screened for relevance to EcR activation and downstream events leading to molting outcomes.
-
Inclusion criteria: mechanistic experimental data linking EcR signaling to endocrine output (ETH/CCAP), neural/motor activity, or apical ecdysis outcomes.
-
Exclusion criteria: studies limited to non-EcR endocrine pathways (unless providing comparative insights), purely descriptive morphology without mechanistic data, or vertebrate-focused studies.
-
-
Quality assessment and evidence integration
-
Evidence for each KE and KER was evaluated using OECD AOP WoE guidance (biological plausibility, essentiality, and empirical support).
-
Comparative studies across insect taxa were considered for domain of applicability.
-
Particular attention was paid to temporal concordance (e.g., timing of EcR activation vs. hormone release vs. motor output), dose–response information, and reproducibility across stressors (genetic manipulation vs. chemical activation).
-
This structured strategy ensured that the AOP is grounded in well-established insect physiology while incorporating toxicological evidence relevant to chemical risk assessment. The documentation of the search scope, terms, and expert scoping facilitates transparency, reproducibility, and re-use of the AOP and its components in other networks addressing arthropod endocrine disruption.
Summary of the AOP
Events:
Molecular Initiating Events (MIE)
Key Events (KE)
Adverse Outcomes (AO)
| Type | Event ID | Title | Short name |
|---|
| MIE | 103 | Increase, Ecdysone receptor hyperactivation | Increase, EcR hyperactivation |
| KE | 2373 | Increase, Mis-timed ecdysone receptor-responsive nuclear receptor cascade | Increase, Mis-timed EcR-responsive NR cascade |
| KE | 988 | Decrease, Circulating ecdysis triggering hormone | Decrease, Circulating ETH |
| KE | 1266 | Decrease, Circulating crustacean cardioactive peptide | Decrease, Circulating CCAP |
| KE | 1267 | Decrease, Ecdysis motor program activity | Decrease, Ecdysis motor program activity |
| KE | 993 | Decrease, Abdominal muscle contraction | Decrease, Abdominal muscle contraction |
| AO | 990 | Increase, Incomplete ecdysis | Increase, Incomplete ecdysis |
| AO | 350 | Increase, Mortality | Increase, Mortality |
Relationships Between Two Key Events (Including MIEs and AOs)
| Title | Adjacency | Evidence | Quantitative Understanding |
|---|
Network View
Prototypical Stressors
Life Stage Applicability
| Life stage | Evidence |
|---|---|
| Adult, reproductively mature | High |
| Juvenile | High |
Taxonomic Applicability
| Term | Scientific Term | Evidence | Link |
|---|---|---|---|
| Arthropoda | Arthropoda | High | NCBI |
Sex Applicability
| Sex | Evidence |
|---|---|
| Unspecific | High |
Overall Assessment of the AOP
The overall weight of evidence (WoE) for this AOP is high, supported by strong biological plausibility, extensive empirical evidence, and moderate but growing quantitative understanding. The mechanistic underpinnings are grounded in well-established endocrine and neurophysiological processes that regulate molting in arthropods.
-
Biological plausibility: The causal linkages between EcR activity, transcriptional cascades, neuropeptide release (ETH, CCAP), motor program activation, and abdominal muscle contraction are strongly supported by fundamental developmental biology. Each KE corresponds to a critical control point in the molting process, and their functional interdependencies are highly conserved across arthropods. The plausibility of the AOP is reinforced by decades of work in insect endocrinology and neurobiology, making the overall pathway mechanistically coherent.
-
Essentiality of key events: Essentiality is considered high. Genetic, pharmacological, and RNAi manipulations at multiple nodes (EcR activity, nuclear receptor genes, ETH/CCAP neurons, motor programs) consistently demonstrate that disruption prevents progression to downstream events and results in incomplete ecdysis. Rescue experiments (e.g., exogenous ETH or CCAP) further confirm causal roles of upstream events in controlling downstream outcomes.
-
Empirical support: Empirical evidence is strong, with consistent observations across multiple model systems, particularly holometabolous insects (Drosophila melanogaster, Manduca sexta, Bombyx mori, Tribolium castaneum). Hyperactivation of EcR by diacylhydrazine insecticides reliably produces predictable phenotypes: altered gene expression, reduced peptide release, suppression of motor activity, incomplete ecdysis, and mortality. Similar outcomes have been observed in crustaceans, though fewer studies exist. The relationships are reproducible and concordant across experimental systems.
-
Quantitative understanding: Current quantitative data are moderate. Some dose–response and temporal dynamics have been characterized for EcR agonists, transcriptional markers, and ETH release, showing concordance between exposure intensity and severity of downstream disruption. However, quantitative thresholds vary by species, and detailed response–response models remain limited. Cross-species extrapolation requires further development.
-
Domain of applicability: The AOP is broadly applicable to arthropods. Evidence is strongest for holometabolous insects, with moderate confidence for crustaceans. Applicability is equal in both sexes and most relevant for juvenile and larval life stages undergoing ecdysis.
Regulatory relevance: This AOP is highly relevant for the assessment of insect growth regulators and other EcR-active chemicals. The robustness of the evidence supports its application in screening, prioritization, test guideline refinement, and ecological risk assessment of non-target arthropods. While quantitative relationships require strengthening, the AOP already provides a reliable mechanistic framework linking receptor-level perturbations to population-relevant adverse outcomes.
Overall conclusion: The AOP is supported by strong mechanistic plausibility, consistent empirical evidence, and partial quantitative understanding. Confidence in the AOP is high for insects and moderate but growing for crustaceans. This AOP is sufficiently developed for regulatory consideration and provides a valuable tool for integrating mechanistic data into chemical hazard and risk assessment.
Domain of Applicability
The applicability of this AOP is defined by the biological role of the ecdysone receptor (EcR) and the conserved endocrine and neuromuscular pathways controlling ecdysis in arthropods. Because these processes are highly conserved, the AOP is broadly relevant across arthropod taxa, though the weight of empirical support differs between groups.
-
Taxonomic applicability:
-
Insects: Evidence is strongest in holometabolous insects (Drosophila melanogaster, Manduca sexta, Bombyx mori, Tribolium castaneum), where EcR function and downstream nuclear receptor cascades have been extensively characterized. Hemimetabolous insects (e.g., locusts, cockroaches) also rely on EcR regulation of molting, although fewer data are available regarding EcR hyperactivation specifically.
-
Crustaceans: Empirical evidence is growing in species such as Daphnia magna and decapods (Carcinus maenas, Penaeus spp.). EcR-mediated endocrine control of molting and neuropeptide signaling (e.g., ETH, CCAP) is conserved, though fewer direct studies of hyperactivation have been conducted.
-
Other arthropods: Arachnids and myriapods also possess EcR-mediated molting control, suggesting applicability; however, direct experimental support for EcR hyperactivation effects is currently limited.
-
-
Sex applicability: The AOP is relevant to both males and females, as EcR signaling and molting are fundamental physiological processes that do not differ between sexes.
-
Life-stage applicability: The pathway is most relevant to larval and juvenile stages undergoing active molting and metamorphic transitions. Hyperactivation of EcR at these stages disrupts the timing of neuropeptide release and muscle activity essential for shedding the old cuticle. In adults, where molting no longer occurs, the AOP is not relevant. In crustaceans, however, periodic molting throughout life stages means applicability extends beyond juveniles.
-
Environmental context: This AOP is applicable in both terrestrial and aquatic environments. Exposure to EcR agonists (e.g., diacylhydrazine insecticides) has been documented in agricultural landscapes affecting insects, and in aquatic systems where run-off or drift may impact crustaceans and non-target insects.
Summary: The AOP is broadly applicable across arthropods, with highest confidence for holometabolous insects and increasing but moderate confidence for crustaceans. It applies equally to both sexes and is particularly relevant for juvenile life stages undergoing ecdysis. Conservation of the underlying molecular and physiological pathways supports the generalization of this AOP across diverse arthropod taxa.
Essentiality of the Key Events
The essentiality of the key events (KEs) in this AOP is strongly supported by genetic, pharmacological, and physiological studies. Experimental manipulations at multiple levels of the pathway demonstrate that blocking or altering a KE prevents downstream events and results in the predicted adverse outcomes.
-
MIE: EcR hyperactivation (Event 103)
-
Evidence: EcR is the master regulator of molting and metamorphosis. Overactivation by non-steroidal agonists (e.g., diacylhydrazines such as tebufenozide, halofenozide) or genetic overexpression leads to inappropriate or prolonged signaling. This produces temporal misalignment of transcriptional cascades and prevents successful molting.
-
Essentiality: High. Experimental induction of EcR hyperactivation consistently produces molting disruption and lethality.
-
-
KE: Mis-timed EcR-responsive nuclear receptor cascade (Event 2373)
-
Evidence: The nuclear receptor cascade (e.g., HR3, E75, Ftz-f1) requires precise temporal induction. EcR hyperactivation disturbs this sequence, leading to abnormal timing or suppression of downstream factors. In Drosophila, blocking or mis-timing expression of these nuclear receptors prevents ecdysis and metamorphosis.
-
Essentiality: High. Temporal mis-regulation of the cascade directly disrupts hormonal and neuronal coordination of ecdysis.
-
-
KE: Decreased circulating ETH (Event 998)
-
Evidence: ETH is secreted by Inka cells and triggers the ecdysis motor program. ETH knockout, RNAi knockdown, or pharmacological suppression prevents ecdysis initiation. Hyperactivation of EcR suppresses normal ETH induction, blocking downstream motor programs. Rescue experiments with ETH peptide partially restore motor activity.
-
Essentiality: High. ETH release is indispensable for activating ecdysis behavior.
-
-
KE: Decreased circulating CCAP (Event 1266)
-
Evidence: CCAP neurons drive motor output and sustain ecdysis behavior. Silencing or ablating CCAP neurons abolishes motor programs. Hyperactivation of EcR alters CCAP timing and suppresses release. Exogenous CCAP application can restore abdominal contractions in some contexts.
-
Essentiality: High. CCAP is required for proper execution of the ecdysis motor program.
-
-
KE: Decreased ecdysis motor program activity (Event 1267)
-
Evidence: Neurophysiological recordings in Manduca sexta and Drosophila show that ETH and CCAP drive motor outputs. Disruption of their release abolishes patterned motor activity. Without the motor program, cuticle shedding cannot occur.
-
Essentiality: High. Blocking motor program activity results directly in incomplete ecdysis.
-
-
KE: Decreased abdominal muscle contraction (Event 993)
-
Evidence: Abdominal contractions generate the mechanical forces required to shed the old cuticle. Pharmacological blockade or neural inhibition of muscle contractions prevents ecdysis. Hyperactivation of EcR suppresses ETH/CCAP signals, reducing contractions.
-
Essentiality: High. Abdominal contractions are indispensable for successful cuticle shedding.
-
-
KE: Incomplete ecdysis (Event 990)
-
Evidence: Incomplete shedding of the old cuticle results in physical entrapment and lethality. This is a consistent and well-documented consequence of endocrine or neuromuscular disruption in molting studies.
-
Essentiality: High. Incomplete ecdysis is the proximate cause of death in affected individuals.
-
-
AO: Mortality (Event 350)
-
Evidence: Mortality is the inevitable outcome of incomplete ecdysis. Laboratory and field studies with EcR agonists demonstrate nearly 100% lethality at effective doses.
-
Overall assessment: The essentiality of all key events in this AOP is rated high. Experimental manipulations at each level (EcR activity, transcriptional cascades, peptide release, motor program activity, muscle contractions) consistently demonstrate that disruption leads to incomplete ecdysis and mortality, while rescue experiments with downstream factors (e.g., ETH, CCAP) can mitigate effects. This provides strong causal evidence that each KE is indispensable for pathway progression.
Evidence Assessment
The evidence supporting the KERs in this AOP is strong and consistent with established knowledge of ecdysteroid signaling, neuropeptide regulation, and neuromuscular control of ecdysis. The assessment below considers biological plausibility, empirical support, and quantitative understanding for each KER.
KER 103 → 2373 (EcR hyperactivation → Mis-timed nuclear receptor cascade)
-
Biological plausibility: Strong. EcR orchestrates temporal induction of nuclear receptors (e.g., E75, HR3, Ftz-f1). Overactivation leads to premature or prolonged expression, disrupting endocrine timing.
-
Empirical support: Exposure of insects to diacylhydrazine agonists (e.g., tebufenozide, halofenozide) produces mis-regulation of early and late response genes, consistent with disrupted transcriptional cascades.
-
Quantitative understanding: Dose–response data exist in Drosophila and Lepidoptera, showing correlation between agonist exposure, transcriptional mis-timing, and molting failure. Cross-species thresholds are less well defined.
KER 2373 → 998 (Mis-timed nuclear receptor cascade → Decreased circulating ETH)
-
Biological plausibility: Moderate to strong. ETH release depends on transcriptional competence established by nuclear receptor signaling. If the cascade is mis-timed, ETH induction fails or occurs at inappropriate developmental stages.
-
Empirical support: ETH secretion is abolished in EcR mutant insects or when nuclear receptor activation is blocked. Hyperactivation by EcR agonists suppresses or delays ETH release in Manduca sexta and Drosophila.
-
Quantitative understanding: Limited. ETH titers decline predictably under EcR hyperactivation, but exact response functions are not established.
KER 2373 → 1266 (Mis-timed nuclear receptor cascade → Decreased circulating CCAP)
-
Biological plausibility: Moderate. CCAP neuron activation is tightly linked to transcriptional control downstream of EcR. Mis-regulated cascades can suppress CCAP release.
-
Empirical support: Electrophysiological and immunohistochemical studies show EcR agonist exposure reduces CCAP release during ecdysis. Knockdown of nuclear receptors upstream of CCAP neurons produces similar results.
-
Quantitative understanding: Sparse. Qualitative evidence is strong, but quantitative measures of CCAP release under EcR hyperactivation remain limited.
KER 998 → 1267 (Decreased ETH → Decreased ecdysis motor program activity)
-
Biological plausibility: Strong. ETH directly activates central pattern generators that drive the ecdysis motor program.
-
Empirical support: ETH knockout or suppression in Drosophila prevents initiation of ecdysis. Exogenous ETH rescues motor activity in mutants and EcR agonist-treated insects.
-
Quantitative understanding: Moderate. Threshold levels of ETH required to activate motor programs are partially characterized in Manduca sexta and Drosophila.
KER 1266 → 1267 (Decreased CCAP → Decreased ecdysis motor program activity)
-
Biological plausibility: Strong. CCAP sustains the motor activity required for cuticle shedding.
-
Empirical support: Silencing CCAP neurons abolishes motor output. Exogenous CCAP restores abdominal contractions in insects exposed to EcR agonists.
-
Quantitative understanding: Limited. While functional restoration has been demonstrated, dose-dependent relationships remain under-characterized.
KER 1267 → 990 (Decreased motor program activity → Incomplete ecdysis)
-
Biological plausibility: Strong. Motor programs are the direct drivers of the physical shedding process. Without sufficient activity, ecdysis fails.
-
Empirical support: Behavioral studies in Manduca sexta and Drosophila show reduced motor program activity correlates with incomplete shedding of the cuticle.
-
Quantitative understanding: Moderate. Time-series data link motor activity suppression with rates of incomplete ecdysis, but quantitative thresholds are not fully defined.
KER 993 → 990 (Decreased abdominal muscle contraction → Incomplete ecdysis)
-
Biological plausibility: Strong. Abdominal contractions generate the force required to rupture and shed the old cuticle.
-
Empirical support: Pharmacological or neurogenic inhibition of abdominal contractions prevents successful molting in insects. EcR agonist treatment reduces contraction frequency and amplitude.
-
Quantitative understanding: Limited. Few studies have established exact contraction thresholds needed for successful ecdysis.
KER 990 → 350 (Incomplete ecdysis → Mortality)
-
Biological plausibility: Strong. Failure to complete ecdysis is incompatible with survival in arthropods.
-
Empirical support: Consistently observed across studies: incomplete molting caused by EcR agonists results in nearly 100% mortality.
-
Quantitative understanding: High at the AO level. Mortality rates scale with frequency of incomplete ecdysis in laboratory exposures, though species sensitivity varies.
Overall WoE Conclusion
-
Biological plausibility: High, based on conserved endocrine and neuromuscular processes across arthropods.
-
Empirical support: Strong, with evidence from multiple insect models and supporting data in crustaceans. Rescue experiments (e.g., exogenous ETH or CCAP) provide compelling causal evidence.
-
Quantitative understanding: Moderate. While qualitative relationships are strong, quantitative dose-response data are incomplete and vary across species.
Confidence in the evidence base for this AOP is high, particularly for insects, with moderate but growing support in crustaceans.
Known Modulating Factors
Several factors are known to modulate the relationship between EcR hyperactivation and the downstream key events leading to incomplete ecdysis and mortality:
-
Developmental stage: Sensitivity is highest immediately prior to and during the molting window (larval–larval molts, larval–pupal transition, and adult emergence). Exposures outside these periods may have limited or no effect.
-
Species/strain differences: Variability in EcR isoform expression, timing of hormone release, or cuticle properties can influence susceptibility across insect taxa. Holometabolous insects appear generally more sensitive to EcR agonists than some hemimetabolous species.
-
Nutritional status: Adequate energy reserves are necessary to sustain neuropeptide release and muscle activity during ecdysis. Nutritionally stressed individuals show increased rates of incomplete ecdysis when challenged with EcR agonists.
-
Environmental conditions: Temperature and humidity modulate the success of molting by influencing hormone secretion kinetics, motor program execution, and cuticle flexibility. For example, lower humidity can exacerbate cuticle hardening and increase mortality when ecdysis is disrupted.
-
Chemical-specific properties: Potency, bioavailability, and persistence of EcR agonists can influence the severity and timing of effects. Lipophilicity and cuticular penetration rates can modulate effective internal concentrations and alter KE progression.
-
Sex: No consistent sex-related differences have been reported; effects are considered sex-independent.
Together, these factors may alter the sensitivity or manifestation of key events, contributing to observed variability across studies and taxa.
| Modulating Factor (MF) | Influence or Outcome | KER(s) involved |
|---|---|---|
Quantitative Understanding
The overall qualitative sequence of events from EcR hyperactivation to mortality is well-supported, but quantitative relationships between individual key events (KERs) are less well defined. Current understanding is as follows:
-
Molecular Initiating Event (EcR hyperactivation): Receptor binding affinity and transcriptional activation thresholds have been characterized for several EcR agonists (e.g., tebufenozide, methoxyfenozide), providing quantitative data at the receptor level. However, translation of receptor occupancy to downstream transcriptional mis-timing remains incompletely modeled.
-
Mis-timed nuclear receptor cascade → Neuropeptide disruption (ETH/CCAP): Temporal concordance is strong; premature or prolonged EcR activation shifts hormone pulse timing. Quantitative dose–response data linking agonist concentration to suppression or delay of ETH/CCAP release are limited, with most studies reporting qualitative or semi-quantitative changes (e.g., altered peak amplitude or delayed onset).
-
Neuropeptide levels → Motor program activity/abdominal contraction: Evidence demonstrates a threshold requirement for ETH and CCAP release to initiate and sustain ecdysis motor programs. Below-threshold neuropeptide levels result in absent or incomplete motor sequences. Some quantitative data exist in Manduca sexta and Drosophila CNS preparations, but standardized metrics (e.g., hormone concentration vs. motor burst frequency) are lacking.
-
Motor activity → Incomplete ecdysis: A largely deterministic relationship. Failure of motor output or reduced abdominal contractions consistently results in incomplete cuticle shedding. This linkage is considered functionally threshold-based rather than graded.
-
Incomplete ecdysis → Mortality: Mortality rates correlate strongly with frequency of incomplete ecdysis events. Quantitative dose–response relationships have been documented in exposure studies with EcR agonists, where increasing chemical concentration yields a proportional increase in incomplete molts and mortality. However, variability in species, life stage, and environmental conditions limits generalization across taxa.
Considerations for Potential Applications of the AOP (optional)
This AOP provides a mechanistic framework that can be applied across several domains of chemical hazard assessment and regulatory decision-making. Because EcR is a critical developmental regulator in arthropods, perturbations at this receptor level are highly predictive of adverse outcomes that impact survival and population sustainability. The structured evidence in this AOP allows for multiple scientific and regulatory applications:
1. Screening and Prioritization of Chemicals
-
The molecular initiating event (EcR hyperactivation) can be evaluated using in vitro receptor-binding assays, reporter gene assays, or in silico modeling (e.g., docking and QSAR approaches).
-
This allows early identification of chemicals with EcR agonist activity, facilitating prioritization for further testing.
2. Development and Refinement of Test Guidelines
-
Intermediate KEs (mis-timed transcriptional cascades, altered neuropeptide release, suppression of motor activity) provide mechanistic biomarkers that could be incorporated into OECD guideline refinements.
-
For example, ETH and CCAP measurements or transcriptomic markers (E75, HR3, Ftz-f1) could be developed into endpoints for short-term assays, reducing reliance on labor-intensive whole-organism tests.
3. Integrated Approaches to Testing and Assessment (IATA)
-
The AOP provides the scientific foundation for integrating multiple data streams (in vitro assays, omics biomarkers, neuropeptide quantification, behavioral endpoints) into tiered testing strategies.
-
Such IATAs can reduce animal use and provide mechanistic clarity for regulatory submissions.
4. Grouping and Read-Across
-
The AOP supports chemical grouping strategies by identifying shared modes of action (EcR agonism).
-
Structural alerts (e.g., diacylhydrazine scaffolds) and activity profiling can be used to apply read-across approaches to new or untested compounds.
5. Ecological Risk Assessment
-
By mechanistically linking molecular initiating events to population-level mortality, this AOP provides a foundation for extrapolating laboratory mechanistic data to ecological risk scenarios.
-
Particularly relevant for non-target insects (e.g., pollinators, natural enemies) and aquatic crustaceans that may be exposed to EcR-active pesticides in agricultural runoff.
-
Population models could incorporate incomplete ecdysis as a measurable endpoint to predict ecological consequences.
6. Regulatory Decision-Making
-
Regulators may use this AOP in weight-of-evidence evaluations for substances with suspected endocrine-disrupting activity in arthropods.
-
It provides a transparent mechanistic rationale that strengthens the scientific basis for restrictions, mitigation measures, or prioritization of monitoring for non-target exposure.
7. Research and Method Development
-
The AOP highlights knowledge gaps in quantitative relationships between KEs, encouraging targeted research.
-
Development of quantitative models (e.g., dose-response functions for ETH and CCAP suppression) could improve predictive utility.
Summary: This AOP has strong potential applications in screening, test guideline refinement, grouping/read-across, and ecological risk assessment. It provides a mechanistic bridge from molecular initiating events to organismal mortality, supporting regulatory decisions concerning the use and approval of EcR-active insecticides and other endocrine-active substances.
References
-
Becker RA, Ankley GT, Edwards SW, et al. Increasing scientific confidence in adverse outcome pathways: Application of tailored Bradford-Hill considerations for evaluating weight of evidence. Toxicol In Vitro. 2015;29:745–53.
-
Clark AC, del Campo ML, Ewer J. Neuroendocrine control of larval ecdysis behavior in Drosophila: Complex coordination of a stereotyped behavior. J Neurosci. 2004;24:4283–92.
-
Dhadialla TS, Carlson GR, Le DP. New insecticides with ecdysteroidal and juvenile hormone activity. Annu Rev Entomol. 1998;43:545–69.
-
Gammie SC, Truman JW. Neuropeptide hierarchies and the activation of sequential motor programs in ecdysis behavior of the moth Manduca sexta. J Neurosci. 1997;17:4389–97.
-
Hormann RE, et al. Superimposition evaluation of ecdysteroid agonist chemotypes using quantitative structure–activity relationships. J Comput Aided Mol Des. 2003;17:319–38.
-
Jindal V, Nagaraju J, Mishra RK. Functional characterization of ecdysis-triggering hormone receptors from Anopheles gambiae. Front Physiol. 2021;12:702979.
-
Kannangara JR, Mirth CK, Warr CG. Regulation of ecdysone production in Drosophila by neuropeptides and peptide hormones. Open Biol. 2021;11:200373.
-
Lahr EC, Dean D, Ewer J. Genetic analysis of ecdysis behavior in Drosophila reveals partially overlapping functions of two unrelated neuropeptides. J Neurosci. 2012;32:6819–29.
-
Lenaerts C, Verlinden H, Vuerinckx K, et al. The ecdysis-triggering hormone system is essential for successful ecdysis in the desert locust, a hemimetabolous insect. Front Physiol. 2017;8:597.
-
Nakagawa Y. Nonsteroidal ecdysone agonists. In: Evans PD, editor. Adv Insect Physiol. Vol. 32. Academic Press; 2005. p. 131–73.
-
OECD. Users’ Handbook Supplement to the Guidance Document for Developing and Assessing Adverse Outcome Pathways. Version 2.7. Paris: OECD Publishing; 2024.
-
Retnakaran A, Krell P, Feng Q, Arif B. Ecdysone agonists: Mechanism and importance in controlling insect pests of agriculture and forestry. Arch Insect Biochem Physiol. 2003;54:187–99.
-
Ruivo R, Frei A, Kümmerer K. The ecdysone receptor case study: Species-specificity of EcR agonists and non-target concerns. Environ Sci Eur. 2021;33:159.
-
Shi Y, Li K, Jia Q, et al. The ecdysis-triggering hormone system, via ETH/ETHR-B signaling, regulates adult reproduction in Bactrocera dorsalis. Front Physiol. 2019;10:151.
-
Song Y, Villeneuve DL, Toyota K, Iguchi T, Tollefsen KE. Ecdysone receptor agonism leading to lethal molting disruption in arthropods: Review and adverse outcome pathway development. Environ Sci Technol. 2017;51(8):4142–57. doi:10.1021/acs.est.7b00480.
-
Sundaram M, et al. Basis for selective action of a synthetic molting hormone agonist, RH-5992 (tebufenozide). Insect Biochem Mol Biol. 1998;28:693–704.
-
Truman JW, Riddiford LM. Endocrine insights into the evolution of metamorphosis in insects. Annu Rev Entomol. 2002;47:467–500.
-
Verbakel L, Lenaerts C, Abou El Asrar R, et al. Prothoracicostatic activity of the ecdysis-regulating neuropeptide crustacean cardioactive peptide (CCAP) in the desert locust. Int J Mol Sci. 2021;22:13465.
-
Wing KD. RH-5849, a nonsteroidal ecdysone agonist: Effects on a Drosophila cell line. Science. 1988;241(4864):467–9.
-
Yamanaka N, Rewitz KF, O’Connor MB. Ecdysone control of developmental transitions: Lessons from Drosophila research. Annu Rev Entomol. 2013;58:497–516.
-
Žitňan D, Adams ME. Excitatory and inhibitory roles of central ganglia in initiation of the insect ecdysis behavioural sequence. J Exp Biol. 2000;203:1329–40.