
This AOP is licensed under the BY-SA license. This license allows reusers to distribute, remix, adapt, and build upon the material in any medium or format, so long as attribution is given to the creator. The license allows for commercial use. If you remix, adapt, or build upon the material, you must license the modified material under identical terms.
AOP: 343
Title
Ecdysone receptor antagonism leading to mortality via mis-timed ecdysone receptor-responsive nuclear receptor cascade
Short name
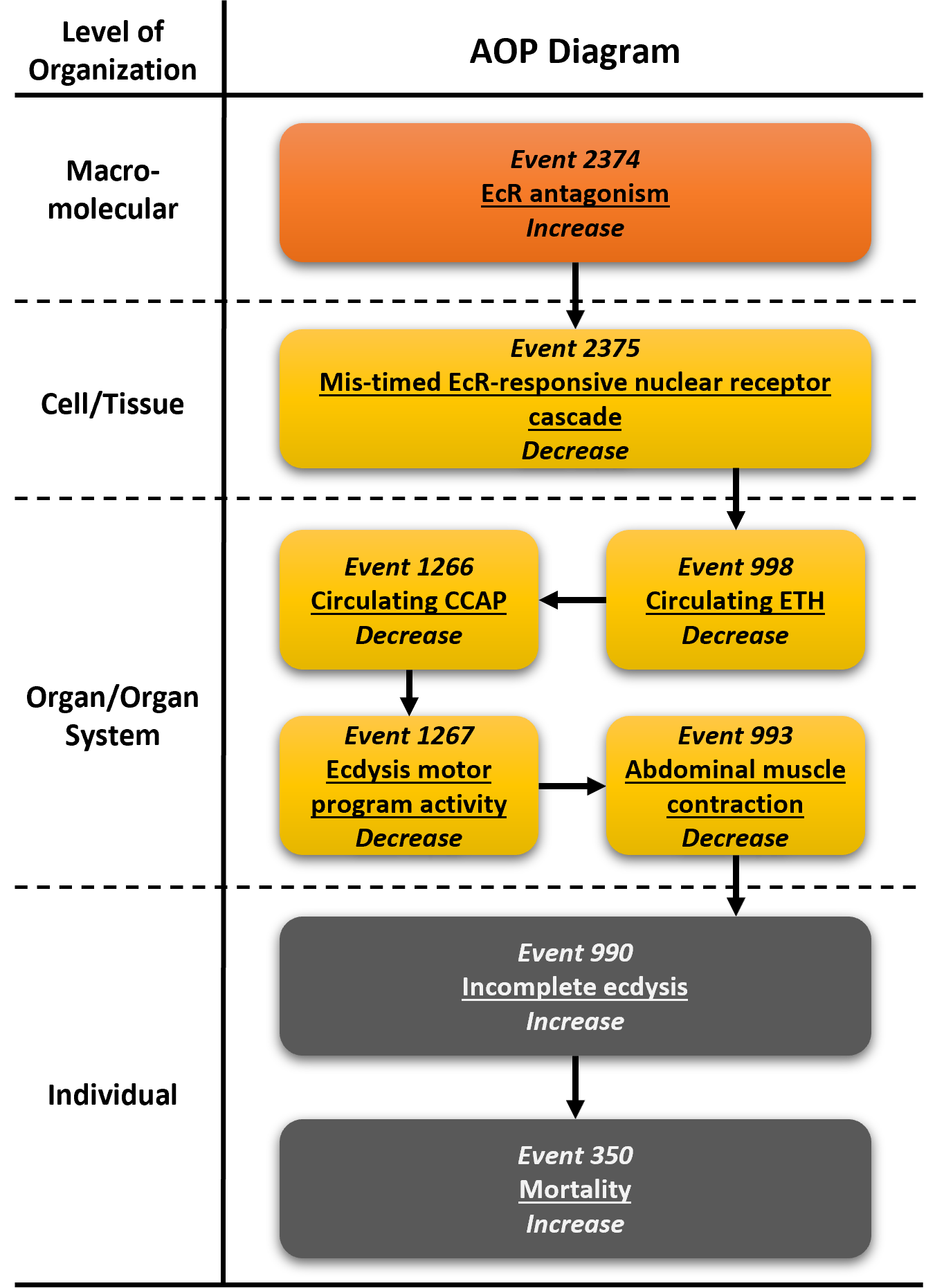
Graphical Representation
Additional AOP Exploration Options
Click links below to explore AOP 343, Ecdysone receptor antagonism leading to mortality via mis-timed ecdysone receptor-responsive nuclear receptor cascade in tools offered by third parties.
Point of Contact
Contributors
- You Song
Coaches
OECD Information Table
| OECD Project # | OECD Status | Reviewer's Reports | Journal-format Article | OECD iLibrary Published Version |
|---|---|---|---|---|
This AOP was last modified on October 01, 2025 16:06
Revision dates for related pages
| Page | Revision Date/Time |
|---|---|
| Increase, Ecdysone receptor antagonism | September 29, 2025 05:17 |
| Decrease, Mis-timed ecdysone receptor-responsive nuclear receptor cascade | September 29, 2025 05:18 |
| Decrease, Circulating ecdysis triggering hormone | May 24, 2018 16:34 |
| Decrease, Circulating crustacean cardioactive peptide | May 24, 2018 16:37 |
| Decrease, Ecdysis motor program activity | September 29, 2025 04:18 |
| Decrease, Abdominal muscle contraction | May 24, 2018 16:41 |
| Increase, Incomplete ecdysis | May 24, 2018 16:41 |
| Increase, Mortality | October 26, 2020 05:18 |
| Increase, EcR antagonism leads to Decrease, Mis-timed EcR-responsive NR cascade | September 29, 2025 05:19 |
| Decrease, Mis-timed EcR-responsive NR cascade leads to Decrease, Circulating ETH | September 29, 2025 05:20 |
| Decrease, Circulating ETH leads to Decrease, Circulating CCAP | February 09, 2017 03:34 |
| Decrease, Circulating CCAP leads to Decrease, Ecdysis motor program activity | February 09, 2017 03:35 |
| Decrease, Ecdysis motor program activity leads to Decrease, Abdominal muscle contraction | September 29, 2025 04:17 |
| Decrease, Abdominal muscle contraction leads to Increase, Incomplete ecdysis | December 03, 2016 16:38 |
| Increase, Incomplete ecdysis leads to Increase, Mortality | December 03, 2016 16:38 |
Abstract
This Adverse Outcome Pathway (AOP) describes how antagonism of the ecdysone receptor (EcR) disrupts endocrine regulation of molting in arthropods, leading to incomplete ecdysis and mortality. EcR normally regulates transcriptional cascades controlling neuropeptide release (ETH, CCAP), motor program activity, and muscle contractions necessary for successful molting. Antagonists block receptor activation, leading to suppression or mis-timing of nuclear receptor signaling, reduced ETH and CCAP release, impaired neuromuscular activity, and ultimately failure to shed the old cuticle. The adverse outcome, mortality, has ecological and regulatory relevance for assessing the effects of environmental contaminants and pesticides with EcR antagonist activity.
AOP Development Strategy
Context
While EcR agonists are widely used as insect growth regulators, EcR antagonists are less common but represent a plausible mode of endocrine disruption. Chemicals acting as antagonists may arise as experimental compounds, metabolites, or environmental contaminants. Understanding the mechanistic links between receptor-level antagonism and organismal outcomes enables prediction of adverse effects in non-target arthropods.
Strategy
The AOP was developed using targeted literature searches in PubMed, Web of Science, and Scopus with keywords including ecdysone receptor antagonism, ecdysis failure, ETH, CCAP, neuropeptides, and arthropod molting disruption. Priority was given to studies using insects (Drosophila melanogaster, Manduca sexta, Tribolium castaneum) and crustaceans (Daphnia magna, Carcinus maenas), where EcR signaling pathways are well characterized. Reviews and OECD guidance on AOP development were used to ensure consistency and regulatory applicability.
Summary of the AOP
Events:
Molecular Initiating Events (MIE)
Key Events (KE)
Adverse Outcomes (AO)
| Type | Event ID | Title | Short name |
|---|
| MIE | 2374 | Increase, Ecdysone receptor antagonism | Increase, EcR antagonism |
| KE | 2375 | Decrease, Mis-timed ecdysone receptor-responsive nuclear receptor cascade | Decrease, Mis-timed EcR-responsive NR cascade |
| KE | 988 | Decrease, Circulating ecdysis triggering hormone | Decrease, Circulating ETH |
| KE | 1266 | Decrease, Circulating crustacean cardioactive peptide | Decrease, Circulating CCAP |
| KE | 1267 | Decrease, Ecdysis motor program activity | Decrease, Ecdysis motor program activity |
| KE | 993 | Decrease, Abdominal muscle contraction | Decrease, Abdominal muscle contraction |
| AO | 990 | Increase, Incomplete ecdysis | Increase, Incomplete ecdysis |
| AO | 350 | Increase, Mortality | Increase, Mortality |
Relationships Between Two Key Events (Including MIEs and AOs)
| Title | Adjacency | Evidence | Quantitative Understanding |
|---|
Network View
Prototypical Stressors
Life Stage Applicability
Taxonomic Applicability
| Term | Scientific Term | Evidence | Link |
|---|---|---|---|
| Arthropoda | Arthropoda | NCBI |
Sex Applicability
Overall Assessment of the AOP
The overall weight of evidence (WoE) supporting this AOP is considered moderate to high, based on strong biological plausibility, a growing body of empirical data, and moderate quantitative understanding. While the majority of evidence derives from insect models, increasing support from crustaceans underscores the broad conservation of EcR signaling in arthropods.
-
Biological plausibility: The pathway is strongly plausible given the central role of EcR in orchestrating transcriptional programs necessary for molting. Antagonism of EcR prevents activation of the nuclear receptor cascade (e.g., HR3, E75, Ftz-f1), which is required for correct timing of neuropeptide release (ETH, CCAP) and motor activity. The link between disrupted receptor signaling and incomplete ecdysis is consistent with fundamental arthropod endocrinology and is highly conserved.
-
Essentiality of the KEs: Essentiality is rated high for all KEs. Blocking EcR directly prevents downstream transcriptional activation. Genetic knockdown or pharmacological suppression of nuclear receptors, ETH, CCAP, or chitin biosynthesis leads to predictable failure of the ecdysis motor program and incomplete shedding. Rescue experiments, such as peptide supplementation, restore motor activity, confirming causal relationships.
-
Empirical support: Empirical evidence is moderate to strong. Insects exposed to EcR antagonists or experimental suppressors show reduced ETH/CCAP secretion, suppressed motor programs, incomplete ecdysis, and mortality. These outcomes are consistent across several insect species (Drosophila melanogaster, Manduca sexta, Tribolium castaneum). Support in crustaceans (Daphnia magna, Carcinus maenas) is emerging but less comprehensive. Results across taxa are concordant and reproducible.
-
Quantitative understanding: Quantitative evidence is limited to moderate. Some dose–response data exist for EcR antagonists, showing relationships between antagonist concentration, suppression of nuclear receptor induction, and failure of molting. However, precise thresholds, variability across species, and predictive cross-species extrapolation remain incompletely characterized. Quantitative modeling of ETH and CCAP suppression and its relationship to ecdysis outcomes is an important data gap.
-
Domain of applicability: Applicability is broad across arthropods. Evidence is strongest for holometabolous insects, moderate for crustaceans, and plausible but not yet tested in other arthropods (arachnids, myriapods). Both sexes are equally susceptible, and the AOP is most relevant for juvenile and larval stages that undergo molting. In crustaceans, the AOP is relevant throughout life due to lifelong molting cycles.
Regulatory relevance: This AOP provides mechanistic support for the screening, prioritization, and assessment of chemicals with potential EcR antagonist activity. It has direct utility for ecological risk assessment of non-target arthropods and could inform refinement of OECD test guidelines by incorporating mechanistic biomarkers (transcriptional markers, neuropeptide measurements). While further quantitative data would improve predictive application, the current evidence base provides sufficient confidence for regulatory consideration.
Overall conclusion: The AOP is supported by strong biological plausibility, high essentiality of KEs, and moderate to strong empirical evidence. Quantitative understanding is developing but not yet comprehensive. Confidence is high for insects and moderate for crustaceans, with potential applicability to other arthropod taxa. This AOP is sufficiently robust to support regulatory application in hazard identification, chemical grouping/read-across, and ecological risk assessment.
Domain of Applicability
The applicability of this AOP is determined by the conservation of ecdysone receptor (EcR) signaling, nuclear receptor cascades, and the endocrine–neuromuscular coordination of molting in arthropods. These processes are essential and broadly conserved, making the AOP relevant to a wide range of taxa, life stages, and ecological contexts.
-
Taxonomic applicability:
-
Insects: Evidence is strongest in holometabolous insects (Drosophila melanogaster, Manduca sexta, Bombyx mori, Tribolium castaneum), where EcR signaling pathways and their role in regulating neuropeptide release and ecdysis are well studied. Hemimetabolous insects (e.g., locusts, cockroaches) also depend on EcR-mediated signaling for molting, though fewer direct data on antagonism are available.
-
Crustaceans: EcR signaling and chitin synthesis are conserved in aquatic crustaceans (Daphnia magna, Carcinus maenas, Penaeus spp.). Several studies demonstrate EcR involvement in molting control, although direct evidence for EcR antagonism is more limited compared to agonism.
-
Other arthropods: Arachnids and myriapods also undergo EcR-regulated molting and chitin synthesis, suggesting the AOP is broadly relevant, although direct empirical data are scarce.
-
-
Sex applicability: The AOP is equally applicable to both sexes. EcR signaling and molting regulation are not sex-dependent, and antagonism of EcR disrupts development and survival irrespective of sex.
-
Life-stage applicability: This AOP is most relevant to juvenile and larval stages that must undergo molting to grow and reach maturity. In insects, applicability is strongest during larval and pupal molts. In crustaceans, molting occurs throughout life, making the AOP relevant across both juvenile and adult stages. Adults in insects that have completed metamorphosis are generally not susceptible, as EcR no longer drives molting processes.
-
Environmental context: The AOP is applicable in both terrestrial and aquatic environments where arthropods are exposed to EcR antagonists. Possible exposure scenarios include:
-
Agricultural systems: pesticide use or residues with unintended EcR antagonist activity.
-
Aquatic ecosystems: runoff or leaching introducing EcR-active contaminants that affect crustaceans and aquatic insects.
-
Laboratory/experimental systems: synthetic EcR antagonists or candidate insecticides tested in model organisms.
-
Summary: This AOP is broadly applicable to arthropods, with high confidence for holometabolous insects, moderate confidence for crustaceans, and lower but plausible confidence for other arthropod groups. It applies to both sexes and is particularly critical during juvenile/larval stages, though in crustaceans it may extend into adulthood due to lifelong molting.
Essentiality of the Key Events
The essentiality of the key events (KEs) in this AOP is considered high overall, as demonstrated by genetic, pharmacological, and physiological evidence. Manipulations at each point of the pathway show that blocking or altering a KE prevents the normal sequence of downstream events, culminating in incomplete ecdysis and mortality.
MIE: EcR antagonism (Event 2374)
-
Evidence: EcR is the master regulator of steroid-induced molting responses. Competitive antagonists, dominant-negative EcR mutations, or EcR knockdowns prevent induction of downstream nuclear receptor genes.
-
Support: In insects, EcR antagonism leads to suppression of HR3, Ftz-f1, and E75 expression, resulting in developmental arrest and failed molting.
-
Essentiality rating: High – antagonism at this level blocks the entire cascade.
KE: Mis-timed EcR-responsive nuclear receptor cascade (Event 2375)
-
Evidence: The cascade of nuclear receptors is essential for establishing endocrine and neuronal competence for molting. If the cascade is not properly activated, subsequent events such as ETH and CCAP release cannot occur at the correct time or magnitude.
-
Support: Knockdown of HR3 or Ftz-f1 in Drosophila prevents ETH induction and ecdysis behavior.
-
Essentiality rating: High – disruption here directly leads to failure of peptide release and downstream events.
KE: Decreased circulating ETH (Event 998)
-
Evidence: ETH released from Inka cells is indispensable for triggering the ecdysis motor program.
-
Support: ETH knockout mutants or RNAi suppression in Drosophila abolish ecdysis behavior. Administration of synthetic ETH rescues ecdysis in some cases, even under EcR disruption.
-
Essentiality rating: High – ETH is the proximate trigger of the motor program.
KE: Decreased circulating CCAP (Event 1266)
-
Evidence: CCAP neurons sustain rhythmic motor outputs during ecdysis. Their activation depends on proper endocrine priming.
-
Support: Ablation or silencing of CCAP neurons abolishes ecdysis motor activity; exogenous CCAP partially restores abdominal contractions. Under EcR antagonism, CCAP release is suppressed.
-
Essentiality rating: High – CCAP is required for maintenance of motor activity during ecdysis.
KE: Decreased ecdysis motor program activity (Event 1267)
-
Evidence: The central pattern generator driving ecdysis behavior must be activated by ETH and maintained by CCAP.
-
Support: Neural recordings show that suppression of ETH/CCAP leads to reduced or absent motor programs. Without this motor program, ecdysis cannot proceed.
-
Essentiality rating: High – the motor program is indispensable for shedding the cuticle.
KE: Decreased abdominal muscle contraction (Event 993)
-
Evidence: Abdominal contractions generate the mechanical forces required for exuviation.
-
Support: Pharmacological suppression of muscle activity or targeted neuromuscular inhibition prevents molting. EcR antagonism reduces contraction frequency and intensity through ETH/CCAP suppression.
-
Essentiality rating: High – abdominal contractions are necessary for successful ecdysis.
KE: Incomplete ecdysis (Event 990)
-
Evidence: Incomplete shedding of the old cuticle results in entrapment and lethality.
-
Support: Consistently observed in EcR antagonist treatments and in peptide or motor program knockdowns.
-
Essentiality rating: High – incomplete ecdysis is the direct cause of mortality.
AO: Mortality (Event 350)
-
Evidence: Mortality is the inevitable outcome of incomplete ecdysis. Individuals unable to shed their old cuticle cannot survive to the next developmental stage.
-
Support: Observed across all insect species tested with EcR antagonists and confirmed in crustaceans under molting disruption conditions.
Overall conclusion on essentiality:
Essentiality of the KEs is high across the entire pathway. Direct manipulations demonstrate that EcR antagonism initiates a cascade of failures in neuropeptide signaling and neuromuscular function, leading inevitably to incomplete ecdysis and death. Rescue studies (e.g., peptide supplementation, gene overexpression) provide further causal evidence that intermediate KEs (ETH, CCAP, motor activity) are indispensable. Confidence in essentiality is highest in insects and moderate but increasing in crustaceans.
Evidence Assessment
The overall evidence for this AOP is supported by strong biological plausibility and increasing empirical support from insect and crustacean models. Each KER is assessed below in terms of biological plausibility, empirical evidence, and quantitative understanding.
KER 2374 → 2375 (EcR antagonism → Mis-timed nuclear receptor cascade)
-
Biological plausibility: Strong. EcR is the central nuclear receptor mediating steroid hormone signaling during molting. Antagonism prevents proper activation of downstream nuclear receptors (e.g., HR3, Ftz-f1, E75).
-
Empirical support: In Drosophila melanogaster and Manduca sexta, EcR knockdowns or exposure to EcR inhibitors suppress nuclear receptor expression, leading to blocked molting.
-
Quantitative understanding: Limited. Qualitative suppression of nuclear receptor targets is clear, but dose–response thresholds for EcR antagonists remain incompletely defined.
KER 2375 → 998 (Mis-timed nuclear receptor cascade → Decreased ETH release)
-
Biological plausibility: Strong. Nuclear receptor signaling is required to establish Inka cell competence for ETH secretion.
-
Empirical support: HR3 and Ftz-f1 knockdown in insects prevents ETH release, resulting in molting failure. In EcR antagonist exposures, ETH levels are reduced or absent.
-
Quantitative understanding: Limited. ETH titer measurements exist for insects exposed to antagonists, but quantitative relationships to receptor inhibition are not fully characterized.
KER 2375 → 1266 (Mis-timed nuclear receptor cascade → Decreased CCAP release)
-
Biological plausibility: Moderate to strong. CCAP neuron activation is downstream of transcriptional and endocrine priming. Antagonism of EcR prevents the cascade needed for their proper activation.
-
Empirical support: Reduced CCAP immunoreactivity and neuronal activity observed in antagonist-treated insects. CCAP knockdown studies independently confirm that reduced CCAP leads to impaired motor programs.
-
Quantitative understanding: Sparse. While CCAP suppression is qualitatively observed, detailed dose–response data are lacking.
KER 998 → 1267 (Decreased ETH → Decreased motor program activity)
-
Biological plausibility: Strong. ETH directly activates central pattern generators in the insect CNS to initiate ecdysis behavior.
-
Empirical support: ETH knockouts in Drosophila abolish motor program activity. Rescue experiments with exogenous ETH restore normal contractions. In EcR antagonist studies, suppression of ETH correlates with reduced or absent motor patterns.
-
Quantitative understanding: Moderate. Threshold ETH concentrations needed for CNS activation have been estimated, but species-specific differences exist.
KER 1266 → 1267 (Decreased CCAP → Decreased motor program activity)
-
Biological plausibility: Strong. CCAP sustains motor activity during ecdysis.
-
Empirical support: Silencing CCAP neurons abolishes rhythmic motor outputs. Exogenous CCAP restores abdominal contractions in insects exposed to EcR modulators.
-
Quantitative understanding: Limited. Data demonstrate binary presence/absence of motor activity but lack dose–response characterization.
KER 1267 → 990 (Decreased motor program activity → Incomplete ecdysis)
-
Biological plausibility: Strong. Coordinated motor programs are required for shedding the old cuticle. Without motor output, the exuvia cannot be shed.
-
Empirical support: In multiple insect species, reduced or absent motor activity directly correlates with incomplete ecdysis. Behavioral recordings show that even partial motor impairment prevents full shedding.
-
Quantitative understanding: Moderate. Time-course and behavioral data are available, but quantitative thresholds for activity loss leading to incomplete ecdysis remain undefined.
KER 993 → 990 (Decreased abdominal muscle contraction → Incomplete ecdysis)
-
Biological plausibility: Strong. Abdominal contractions are the physical mechanism for exuviation.
-
Empirical support: Pharmacological or neural inhibition of muscle contractions leads to incomplete molting in Manduca sexta and Drosophila. In EcR antagonist treatments, contraction amplitude and frequency are suppressed.
-
Quantitative understanding: Limited. Data are mostly descriptive, with few quantitative models linking contraction deficits to molting failure.
KER 990 → 350 (Incomplete ecdysis → Mortality)
-
Biological plausibility: Strong. Failure to shed the cuticle is universally lethal, as the animal remains trapped in the old exoskeleton.
-
Empirical support: In both insects and crustaceans, incomplete molting caused by EcR disruption results in nearly 100% mortality.
-
Quantitative understanding: High at the AO level; survival is binary (ecdysis complete = survival, incomplete = death).
Overall WoE conclusion:
-
Biological plausibility: High, with strong support across all KERs.
-
Empirical support: Moderate to strong. Evidence is strongest in insects, with emerging but more limited support in crustaceans.
-
Quantitative understanding: Moderate. While qualitative evidence is robust, quantitative dose–response data are incomplete, and cross-species extrapolation remains uncertain.
Confidence in the overall AOP: High for insects (Diptera, Lepidoptera, Coleoptera); moderate for crustaceans; plausible for other arthropods but requiring additional data.
Known Modulating Factors
Several intrinsic and extrinsic factors may modulate the progression of this AOP by altering the strength, timing, or outcome of EcR antagonism and downstream key events. While the core pathway is highly conserved and robust, these factors can influence susceptibility across taxa, life stages, and environmental conditions.
-
Temperature: Molting is a temperature-dependent process. Higher or lower temperatures alter ecdysteroid metabolism and the kinetics of receptor activation. In antagonism scenarios, suboptimal temperatures may exacerbate delays or failures in nuclear receptor activation and peptide release, accelerating ecdysis failure.
-
Nutritional status: Adequate energy and nutrient reserves are critical for steroid hormone synthesis, chitin deposition, and neuromuscular activity. Nutrient deprivation can lower circulating ecdysteroid levels, thereby amplifying the impact of EcR antagonism and increasing the likelihood of molting disruption.
-
Developmental stage: Sensitivity to EcR antagonists is greatest during late larval or juvenile stages, when molting cycles are most frequent and tightly regulated. Earlier instars may be more resilient due to developmental plasticity, while late instars and pupal stages are highly vulnerable because mistimed or incomplete molts are often lethal.
-
Species-specific sensitivity: Insects and crustaceans vary in their sensitivity to EcR antagonists. For example, lepidopterans and coleopterans show higher sensitivity than dipterans in some studies, while in crustaceans, species with rapid molting cycles (e.g., Daphnia magna) may be more affected than those with longer intervals between molts. These differences may relate to EcR isoform diversity, receptor binding affinity, and compensatory mechanisms.
-
Ecological and environmental stressors: Stressors such as hypoxia, salinity fluctuations, or exposure to additional contaminants (e.g., pesticides, heavy metals) can compound the effects of EcR antagonism. For instance, stress-induced endocrine disruption may synergize with receptor antagonism to further suppress ETH or CCAP release.
-
Chemical-specific properties: Potency, receptor binding affinity, and bioavailability of EcR antagonists vary widely. Lipophilicity, stability in the environment, and metabolic degradation influence effective concentrations at the receptor level. Weak antagonists may only disrupt molting under conditions of co-exposure to other stressors.
-
Sex and reproductive status: Although molting itself is not sex-specific, reproductive status may modulate endocrine activity in crustaceans. For example, females undergoing vitellogenesis may have altered hormone dynamics that could interact with EcR antagonism.
Summary: The primary modulating factors influencing this AOP include temperature, nutrition, developmental stage, and species-specific physiology. Environmental co-stressors and chemical-specific potency also modify outcomes. While the AOP is mechanistically robust, these factors may shift the dose–response relationships, timing, or severity of adverse outcomes and should be considered in ecological risk assessment frameworks.
| Modulating Factor (MF) | Influence or Outcome | KER(s) involved |
|---|---|---|
Quantitative Understanding
Quantitative understanding of this AOP is moderate, with several well-described dose–response relationships for EcR antagonists in insects, but limited cross-species data and few predictive models that cover the entire cascade. The available evidence supports the directionality and causal links between KEs, but precise thresholds, dynamic ranges, and interspecies extrapolation remain uncertain.
-
Molecular initiating event (EcR antagonism): In vitro receptor-binding and reporter assays have been used to determine antagonist potency (IC₅₀, Ki values) for specific compounds. Competitive inhibition assays demonstrate dose-dependent suppression of EcR activation, but interspecies differences in receptor isoforms complicate extrapolation.
-
EcR-responsive nuclear receptor cascade: Transcriptomic studies in Drosophila melanogaster and Manduca sexta have shown that antagonists reduce or delay expression of nuclear receptor targets (e.g., HR3, Ftz-f1, E75) in a concentration-dependent manner. While these data establish concordance, quantitative thresholds vary widely by chemical and species.
-
Neuropeptide levels (ETH, CCAP): Dose-dependent decreases in ETH and CCAP release have been observed under EcR antagonism. ETH titers in hemolymph decline significantly at antagonist concentrations above established IC₅₀ values. However, quantitative models linking peptide suppression to motor program failure are incomplete, and available data are primarily descriptive.
-
Motor program activity and abdominal contractions: Electrophysiological and behavioral assays reveal that reductions in ETH/CCAP correspond to impaired or absent motor activity. Some dose–response data exist, showing stepwise decreases in contraction frequency and amplitude with antagonist exposure. However, thresholds at which contractions become insufficient for complete molting are not fully defined.
-
Incomplete ecdysis and mortality: Laboratory exposures to EcR antagonists show a clear dose-dependent increase in incomplete molting, with mortality approaching 100% at high concentrations. Survival curves for insects exposed to different antagonist doses show steep responses, indicating limited tolerance margins. These data provide the strongest quantitative linkage in the pathway.
-
Cross-species extrapolation: Data are most developed for holometabolous insects. Crustacean studies (e.g., Daphnia magna) report delayed or failed molts under antagonist exposure, but quantitative ETH/CCAP measurements are lacking. Differences in life-history traits (e.g., molting frequency, hormonal dynamics) complicate extrapolation between taxa.
-
Uncertainties and data gaps:
-
Lack of standardized assays for quantifying ETH and CCAP suppression across species.
-
Few dynamic or mathematical models describing response–response relationships between KEs.
-
Limited quantitative data for crustaceans, arachnids, and other non-insect arthropods.
-
Species-specific EcR isoforms may show variable sensitivity to antagonists, complicating generalization.
-
Summary: Quantitative relationships are best defined at the extremes of the pathway—at the MIE (receptor binding/activation assays) and the AO (dose-dependent mortality). Intermediate KEs (nuclear receptor expression, peptide release, motor activity) are supported by concentration-dependent observations but lack robust dose–response models. The overall confidence in quantitative understanding is moderate for insects, low to moderate for crustaceans, and uncertain for other arthropods. Developing standardized biomarkers and quantitative models would significantly improve predictive application of this AOP.
Considerations for Potential Applications of the AOP (optional)
This AOP offers a mechanistic framework for evaluating how EcR antagonists can disrupt molting and survival in arthropods. Its structured evidence allows regulatory, research, and industry stakeholders to apply the pathway in multiple contexts, from early hazard identification to ecological risk assessment.
1. Screening and Prioritization of Chemicals
-
The molecular initiating event (EcR antagonism) can be probed with in vitro assays (e.g., competitive binding, receptor transactivation inhibition) or in silico tools such as docking models and QSARs.
-
Such assays provide early alerts for potential EcR antagonist activity, enabling regulators to prioritize chemicals for further testing before in vivo exposures occur.
2. Development and Refinement of Test Guidelines
-
This AOP identifies intermediate KEs—nuclear receptor suppression, ETH and CCAP release, motor program activity—that can serve as mechanistic biomarkers.
-
These biomarkers could be incorporated into OECD test guidelines or used as endpoints in short-term assays, reducing reliance on lengthy whole-organism tests.
-
For example, hemolymph peptide quantification or transcriptomic assays targeting HR3 and Ftz-f1 could be integrated into guideline refinements.
3. Integrated Approaches to Testing and Assessment (IATA)
-
The pathway supports IATA by allowing multiple data streams (in vitro binding, gene expression, peptide quantification, motor activity assays) to be combined into tiered testing strategies.
-
This enables more efficient, mechanistically informed decision-making and helps minimize animal use.
4. Grouping and Read-Across
-
EcR antagonists often share structural or functional features. This AOP supports grouping approaches by defining the mechanistic basis of their action.
-
Read-across frameworks can be applied where one antagonist is well-studied, allowing predictions for structurally related chemicals.
-
QSARs built on antagonist scaffolds could provide predictive capability for new or untested substances.
5. Ecological Risk Assessment
-
The AOP provides a mechanistic bridge from molecular receptor-level disruption to population-relevant adverse outcomes.
-
Particularly relevant for non-target insects (e.g., pollinators, natural enemies in agriculture) and aquatic crustaceans exposed via runoff or wastewater discharges.
-
Molting failure can be directly integrated into population models as a measurable endpoint to estimate effects on population dynamics and ecosystem stability.
6. Regulatory Decision-Making
-
Regulators can apply this AOP in weight-of-evidence evaluations of suspected endocrine-active substances.
-
It provides mechanistic support for identifying and managing risks from EcR antagonists in regulatory dossiers, including hazard classification, restriction decisions, and monitoring priorities.
-
By framing mechanistic links transparently, the AOP helps align regulatory actions with the OECD guidance on endocrine disruption.
7. Research and Method Development
-
This AOP highlights several knowledge gaps, particularly in quantitative relationships between intermediate KEs and across species.
-
It encourages the development of standardized assays for ETH/CCAP measurement and motor activity quantification.
-
Further research could also focus on crustacean models, where molting disruption is ecologically critical but empirical data remain limited.
Summary: This AOP can be applied to chemical screening, grouping/read-across, guideline refinement, and ecological risk assessment. It also provides a framework for developing IATA and supports regulatory decision-making on endocrine disruptors. By linking receptor-level antagonism to mortality, the AOP enhances the scientific foundation for evaluating chemical safety in non-target arthropods.