
This AOP is licensed under the BY-SA license. This license allows reusers to distribute, remix, adapt, and build upon the material in any medium or format, so long as attribution is given to the creator. The license allows for commercial use. If you remix, adapt, or build upon the material, you must license the modified material under identical terms.
AOP: 571
Title
5α-reductase inhibition leading to hypospadias in male (mammalian) offspring
Short name
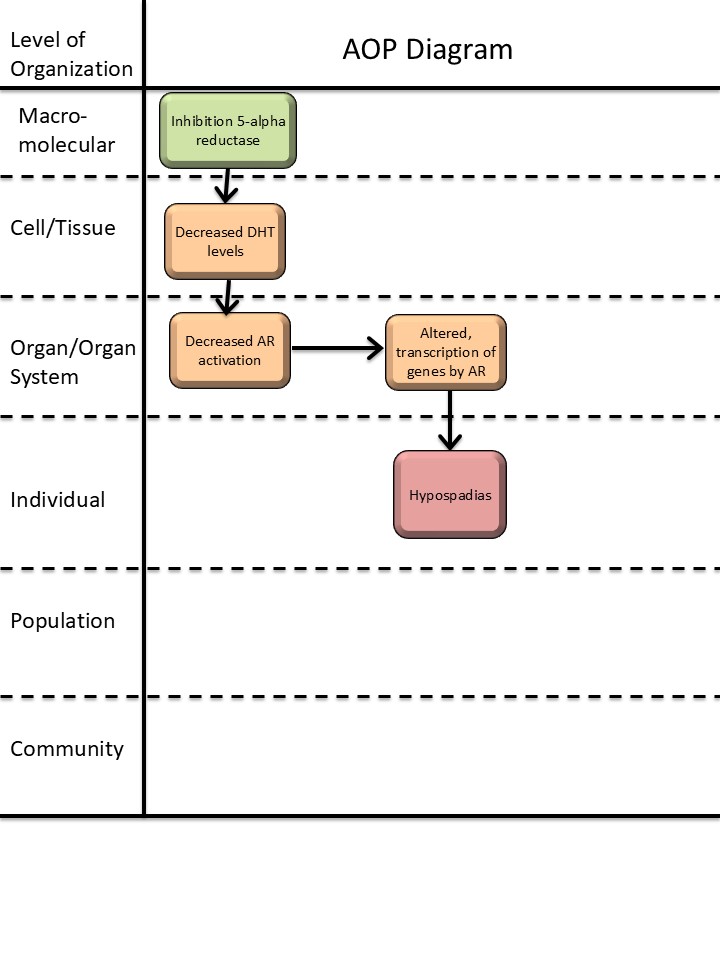
Graphical Representation
Additional AOP Exploration Options
Click links below to explore AOP 571, 5α-reductase inhibition leading to hypospadias in male (mammalian) offspring in tools offered by third parties.
Point of Contact
Contributors
- Terje Svingen
- Emilie Elmelund
Coaches
OECD Information Table
| OECD Project # | OECD Status | Reviewer's Reports | Journal-format Article | OECD iLibrary Published Version |
|---|---|---|---|---|
| Scientific Review |
This AOP was last modified on March 20, 2026 08:39
Revision dates for related pages
| Page | Revision Date/Time |
|---|---|
| Inhibition, 5α-reductase | March 20, 2026 11:45 |
| Decrease, dihydrotestosterone (DHT) levels | March 20, 2026 11:52 |
| Decrease, androgen receptor activation | March 20, 2026 11:54 |
| Altered, Transcription of genes by the androgen receptor | March 20, 2026 12:02 |
| Hypospadias, increased | March 20, 2026 06:37 |
| Inhibition, 5α-reductase leads to Decrease, DHT level | March 20, 2026 12:20 |
| Decrease, AR activation leads to Hypospadias | March 20, 2026 05:58 |
| Decrease, DHT level leads to Decrease, AR activation | March 20, 2026 12:41 |
| Decrease, AR activation leads to Altered, Transcription of genes by the AR | March 20, 2026 10:30 |
| Finasteride | November 29, 2016 18:42 |
Abstract
This AOP links in utero inhibition of 5α-reductase with hypospadias in male offspring. Hypospadias is a common reproductive disorder with a prevalence of up to ~1/125 newborn boys, though with high inter-country variability (Leunbach et al., 2025; Paulozzi, 1999). Developmental exposure to endocrine disrupting chemicals is suspected to contribute to some cases of hypospadias (Mattiske & Pask, 2021). Hypospadias can be indicative of fetal disruptions to male reproductive development, and is associated with short anogenital distance and cryptorchidism (Skakkebaek et al., 2016). Thus, hypospadias is included as a genital abnormality in OECD test guidelines (TG) for developmental and reproductive toxicity (TG 414, 416, 421/422, and 443; (OECD, 2016b, 2016a, 2018a, 2018b, 2021)), as both a measurement of adverse reproductive effects and a direct clinical adverse outcome.
5α-reductase is an enzyme that converts testosterone to dihydrotestosterone (DHT). In normal male reproductive development, DHT activates the androgen receptor (AR) in peripheral reproductive tissues to drive differentiation of the male phenotype, including development of the penis. While testosterone also acts directly at the AR, DHT is 5-10 times more potent, and in peripheral tissues, conversion to DHT is necessary for proper masculinization (Amato et al., 2022; Davey & Grossmann, 2016). This AOP delineates the evidence that inhibition of 5α-reductase reduces DHT levels and consequently AR activation, thereby disrupting penis development and causing hypospadias. The AOP is supported by in vitro experiments upstream of AR activation and by in vivo and human case studies downstream of AR activation. Downstream of a reduction in AR activation, the molecular mechanisms of hypospadias development are not fully delineated, highlighting a knowledge gap in this AOP. Thus, the AOP has potential for inclusion of additional KEs and elaboration of molecular causality links, once these are established. Given that hypospadias is both a clinical and toxicological endpoint, this AOP is considered highly relevant in a regulatory context.
AOP Development Strategy
Context
This AOP is a part of an AOP network for reduced androgen receptor activation causing hypospadias in male offspring. The other AOPs in this network are AOP-477 (‘Androgen receptor antagonism leading to hypospadias in male (mammalian) offspring’), and AOP-570 (‘Decreased testosterone synthesis leading to hypospadias in male (mammalian) offspring’). The purpose of the AOP network is to organize the well-established evidence for anti-androgenic mechanisms-of-action leading to hypospadias, thus informing predictive toxicology and identifying knowledge gaps for investigation and method development.
This work received funding from the European Food and Safety Authority (EFSA) under Grant agreement no. GP/EFSA/PREV/2022/01 and from the Danish Environmental Protection Agency under the Danish Center for Endocrine Disrupters (CeHoS).
Strategy
The OECD AOP Developer’s Handbook was followed alongside pragmatic approaches (Svingen et al., 2021).
KEs and upstream KER1880 (‘inhibition, 5α-reductase leads to decrease, DHT levels’), KER-1935 (‘decrease, DHT levels leads to decrease, AR activation’), and KER-2124 (‘decrease, AR activation leads to altered, transcription of genes by AR’) were considered canonical knowledge and part of an upstream anti-androgenic network developed using mainly key review articles (Draskau et al., 2024). The non-adjacent KER-2828 linking reduced AR activation with hypospadias was developed using a systematic weight-of-evidence approach, following methodology outlined in (Holmer et al., 2024). Articles were retrieved by literature searches in PubMed and Web of Science and extensive screening using pre-defined inclusion and exclusion criteria. Evaluation of methodological reliability of in vivo animal studies was performed using the Science in Risk Assessment and Policy (SciRAP) online tool. As there are currently no in vivo methods to measure AR activation in mammals, six chemicals with known anti-androgenic mechanisms-of-action were chosen for the empirical evidence for this KER. To supplement the in vivo toxicity studies, human case studies and epidemiologic studies were included in the KER. These studies were not systematically evaluated for reliability but served as supporting evidence.
Regarding the inclusion of KEs and KERs, the rationale for the upstream anti-androgenic network is detailed in (Draskau et al., 2024). The link between the upstream network, more specifically KE-286 (‘altered, transcription of genes by AR’), and AO-2082 (‘hypospadias’) likely contains a tissue-specific KE that has not been developed, as sufficient evidence is not yet available. Thus currently, the strongest evidence linking the upstream anti-androgenic network to hypospadias is captured by KER-2828.
Summary of the AOP
Events:
Molecular Initiating Events (MIE)
Key Events (KE)
Adverse Outcomes (AO)
| Type | Event ID | Title | Short name |
|---|
| MIE | 1617 | Inhibition, 5α-reductase | Inhibition, 5α-reductase |
| KE | 1613 | Decrease, dihydrotestosterone (DHT) levels | Decrease, DHT level |
| KE | 1614 | Decrease, androgen receptor activation | Decrease, AR activation |
| KE | 286 | Altered, Transcription of genes by the androgen receptor | Altered, Transcription of genes by the AR |
| AO | 2082 | Hypospadias, increased | Hypospadias |
Relationships Between Two Key Events (Including MIEs and AOs)
| Title | Adjacency | Evidence | Quantitative Understanding |
|---|
| Decrease, AR activation leads to Hypospadias | non-adjacent | High |
Network View
Prototypical Stressors
| Name |
|---|
| Finasteride |
Life Stage Applicability
| Life stage | Evidence |
|---|---|
| Foetal | High |
Taxonomic Applicability
Sex Applicability
| Sex | Evidence |
|---|---|
| Male | High |
Overall Assessment of the AOP
Domain of Applicability
Although the upstream part of the AOPN has a broad applicability domain, the overall AOPN is considered only applicable to male mammals during fetal life, restricted by the applicability of KER-2828 (‘Decrease, AR activation leads to hypospadias’). The term hypospadias is mainly used for describing malformation of the male, and not female, external genitalia. Some studies refer to hypospadias in females, but these have not been reported to be caused by exposure to 5α-reductase inhibitors, and the mechanisms behind these malformations are likely different from the mechanisms in males (Greene, 1937; Stewart et al., 2018). The genital tubercle is programmed by androgens to differentiate into a penis in fetal life in the masculinization programming window, followed by the morphologic differentiation (Welsh et al., 2008). In humans, hypospadias is diagnosed at birth and can also often be observed in rats and mice at this time point, although the rodent penis does not finish developing until a few weeks after birth (Baskin & Ebbers, 2006; Sinclair et al., 2017). The disruption to androgen programming leading to hypospadias thus take place during fetal life, but the AO is best detected postnatally. Specifically, the masculinization programming window (~gestational days (GD) 16-20 in rat, presumably gestational weeks (GW)8-14 in humans) is the primary fetal window of applicability, but effects outside of this window in fetal life, after androgen production has started, cannot be excluded. Regarding taxonomic applicability, hypospadias has mainly been identified in rodents and humans, and the evidence in this AOP is almost exclusively from these species. It is, however, biologically plausible that the AOP is applicable to other mammals as well, given the conserved role of androgens in mammalian reproductive development, and hypospadias has been observed in many domestic animal and wildlife species, albeit not coupled to 5α-reductase inhibition.
Essentiality of the Key Events
|
Event |
Evidence |
Uncertainties and inconsistencies |
|
MIE-1617 Inhibition, 5α-reductase (high) |
Biological plausibility provides strong support for the essentiality of this event, as DHT (produced by 5α-reductase) is one of the primary drivers of penis development. In utero exposure to the 5α-reductase inhibitor finasteride can cause hypospadias in male rats (Clark et al., 1993) Human case studies of 5α-reductase deficiency support the essentiality of this KE, as mutations in 5α-reductase can cause low DHT levels and associated hypospadias in males (Robitaille & Langlois, 2020). See also Table 4 in KER-2828 listing disruptions of AR activity associated with hypospadias in humans. |
In the human case studies, DHT is only measured postnatally and not in fetal life. |
|
KE-1613 Decrease, DHT levels (moderate) |
Biological plausibility provides strong support for the essentiality of this event, as DHT is a ligand of the AR and one of the primary drivers of penis development. Indirect evidence of impact of decreased DHT levels (KE-1613) on AR activation in vivo (KE-1614) comes from the fact that androgen deprivation is used as treatment for prostate cancer, including 5α-reductase inhibitors, to reduce DHT levels and cancer growth (Aggarwal et al., 2010) Moreover, indirect evidence of impact of decreased DHT levels (KE-1613) on AR activation in vitro comes from increasing concentrations of DHT leading to increasing AR activation in vitro in AR reporter gene assays (OECD, 2023; Williams et al., 2017) In patients with 5α-reductase deficiency, DHT levels are reduced and hypospadias is frequently observed, as listed in Table 4 in KER-2828. |
In the human case studies, DHT is only measured postnatally and not in fetal life, As hypospadias is a congenital malformation, it cannot be “reversed” by postnatal DHT treatment. |
|
KE-1614 Decrease, AR activation (moderate) |
Biological plausibility provides strong support for the essentiality of this event, as AR activation is critical for normal penis development.
Conditional or full knockout of Ar in mice results in partly or full sex-reversal of males, including a female-like urethral opening (Willingham et al., 2006; Yucel et al., 2004; Zheng et al., 2015). Human subjects with AR mutations may also have associated hypospadias (as listed in Table 4 in KER-2828). |
|
|
KE-286 Altered, transcription of genes by AR (moderate) |
Biological plausibility provides support for the essentiality of this event. AR is a nuclear receptor and transcription factor regulating transcription of genes, and androgens, acting through AR, are essential for normal male penis development. Genomic AR signaling is necessary and possibly sufficient to drive penis differentiation (Cunha et al., 2021) Known AR-responsive genes active in normal penis development have been thoroughly reviewed (Amato et al., 2022). |
There are currently no AR-responsive genes proved to be causally involved in hypospadias, and it is known that the AR can also signal through non-genomic actions, though these alone are not sufficient for penis differentiation –and genomic AR signaling is necessary for penis development (Cunha et al., 2021; Leung & Sadar, 2017). |
|
Event |
Direct evidence |
Indirect evidence |
Contradictory evidence |
Overall essentiality assessment |
|
MIE 1617 |
*** |
|
|
High |
|
KE 1613 |
* |
** |
|
Moderate |
|
KE 1614 |
** |
|
|
Moderate |
|
KE 286 |
** |
* |
|
Moderate |
Evidence Assessment
The confidence in each of the KERs comprising the AOP are judged as high, with both high biological plausibility and high confidence in the empirical evidence. The mechanistic link between KE-286 (‘altered, transcription of genes by AR’) and AO-2082 (‘hypospadias’) is not established, but given the high confidence in the KERs including the non-adjacent KER-2828 linking to the AO, the overall confidence in the AOP is judged as high.
|
KER |
Biological Plausibility |
Empirical Evidence |
Rationale |
|
KER-1880 Inhibition, 5α-reductase leads to decrease, DHT levels |
High |
High (canonical) |
It is well established that 5α-reductase converts testosterone to DHT. In vitro, in vivo and human studies with 5α-reductase inhibitors have shown dose-dependent decreases in formation of DHT (Draskau et al., 2024). |
|
KER-1935 Decrease, DHT levels leads to decrease, AR activation |
High |
High (canonical) |
It is well established that DHT activates the AR. Direct evidence for this KER is not possible since KE-1614 can currently not be measured and is considered an in vivo effect. Indirect evidence using proxy read-outs of AR activation, either in vitro or in vivo strongly supports the relationship (Draskau et al., 2024). |
|
KER-2124 Decrease, AR activation leads to altered, transcription of genes by AR |
High |
High (canonical) |
It is well established that the AR regulates gene transcription. In vivo animal studies and human genomic profiling show tissue-specific changes to gene expression upon disruption of AR (Draskau et al., 2024). |
|
KER-2828 Decrease, AR activation leads to hypospadias |
High |
High |
It is well established that AR drives penis differentiation. Numerous in vivo toxicity studies and human case studies indirectly show that decreased AR activation leads to hypospadias, with few inconsistencies. The empirical evidence moderately supports temporal concordance for the KER. It should be recognized that the upstream KE-1614 cannot currently be measured directly (in vivo). Instead, empirical evidence was therefore collected for substances known to affect upstream events. This limitation is not considered to lower the strength of the evidence in this case. |
Known Modulating Factors
|
Modulating factor (MF) |
Influence or Outcome |
KER(s) involved |
|
Genotype |
Extended CAG repeat length in AR is associated with reduced AR activity (Chamberlain et al., 1994; Tut et al., 1997). This MF could initiate the AOP at lower stressor doses. |
KER-1935, KER-2124, KER-2828 |
|
Androgen deficiency syndrome |
Low circulating testosterone levels due to hypogonadism (Bhasin et al., 2010). This MF could lower availability of testosterone for conversion by 5α-reductase and thus initiate the AOP at lower stressor doses. |
KER-1935 |
Quantitative Understanding
The quantitative understanding of this AOP is judged as low.
Considerations for Potential Applications of the AOP (optional)
References
Aggarwal, S., Thareja, S., Verma, A., Bhardwaj, T. R., & Kumar, M. (2010). An overview on 5α-reductase inhibitors. Steroids, 75(2), 109–153. https://doi.org/10.1016/j.steroids.2009.10.005
Amato, C. M., Yao, H. H.-C., & Zhao, F. (2022). One Tool for Many Jobs: Divergent and Conserved Actions of Androgen Signaling in Male Internal Reproductive Tract and External Genitalia. Frontiers in Endocrinology, 13, 910964. https://doi.org/10.3389/fendo.2022.910964
Baskin, L., & Ebbers, M. (2006). Hypospadias: Anatomy, etiology, and technique. Journal of Pediatric Surgery, 41(3), 463–472. https://doi.org/10.1016/j.jpedsurg.2005.11.059
Bhasin, S., Cunningham, G. R., Hayes, F. J., Matsumoto, A. M., Snyder, P. J., Swerdloff, R. S., & Montori, V. M. (2010). Testosterone Therapy in Men with Androgen Deficiency Syndromes: An Endocrine Society Clinical Practice Guideline. The Journal of Clinical Endocrinology & Metabolism, 95(6), 2536–2559. https://doi.org/10.1210/jc.2009-2354
Chamberlain, N. L., Driver, E. D., & Miesfeld, R. L. (1994). The length and location of CAG trinucleotide repeats in the androgen receptor N-terminal domain affect transactivation function. Nucleic Acids Research, 22(15), 3181–3186. https://doi.org/10.1093/nar/22.15.3181
Clark, R. L., Anderson, C. A., Prahalada, S., Robertson, R. T., Lochry, E. A., Leonard, Y. M., Stevens, J. L., & Hoberman, A. M. (1993). Critical Developmental Periods for Effects on Male Rat Genitalia Induced by Finasteride, a 5α-Reductase Inhibitor. Toxicology and Applied Pharmacology, 119(1), 34–40. https://doi.org/10.1006/taap.1993.1041
Cunha, G. R., Cao, M., Derpinghaus, A., Baskin, L. S., Cooke, P., & Walker, W. (2021). Cornification and classical versus nonclassical androgen receptor signaling in mouse penile/preputial development. Differentiation, 121, 1–12. https://doi.org/10.1016/j.diff.2021.08.002
Davey, R. A., & Grossmann, M. (2016). Androgen Receptor Structure, Function and Biology: From Bench to Bedside. The Clinical Biochemist. Reviews, 37(1), 3–15.
Draskau, M., Rosenmai, A., Bouftas, N., Johansson, H., Panagiotou, E., Holmer, M., Elmelund, E., Zilliacus, J., Beronius, A., Damdimopoulou, P., van Duursen, M., & Svingen, T. (2024). Aop Report: An Upstream Network for Reduced Androgen Signalling Leading to Altered Gene Expression of Ar Responsive Genes in Target Tissues. Environ Toxicol Chem, (In Press).
Greene, R. R. (1937). Production of Experimental Hypospadias in the Female Rat. Proceedings of the Society for Experimental Biology and Medicine, 36(4), 503–506. https://doi.org/10.3181/00379727-36-9287P
Holmer, M. L., Zilliacus, J., Draskau, M. K., Hlisníková, H., Beronius, A., & Svingen, T. (2024). Methodology for developing data-rich Key Event Relationships for Adverse Outcome Pathways exemplified by linking decreased androgen receptor activity with decreased anogenital distance. Reproductive Toxicology, 128, 108662. https://doi.org/10.1016/j.reprotox.2024.108662
Leunbach, T. L., Berglund, A., Ernst, A., Hvistendahl, G. M., Rawashdeh, Y. F., & Gravholt, C. H. (2025). Prevalence, Incidence, and Age at Diagnosis of Boys With Hypospadias: A Nationwide Population-Based Epidemiological Study. Journal of Urology, 213(3), 350–360. https://doi.org/10.1097/JU.0000000000004319
Leung, J. K., & Sadar, M. D. (2017). Non-Genomic Actions of the Androgen Receptor in Prostate Cancer. Frontiers in Endocrinology, 8. https://doi.org/10.3389/fendo.2017.00002
Mattiske, D. M., & Pask, A. J. (2021). Endocrine disrupting chemicals in the pathogenesis of hypospadias; developmental and toxicological perspectives. Current Research in Toxicology, 2, 179–191. https://doi.org/10.1016/j.crtox.2021.03.004
OECD. (2001). Test No. 416: Two-Generation Reproduction Toxicity [OECD Guidelines for the Testing of Chemicals, Section 4]. OECD Publishing. https://doi.org/10.1787/9789264070868-en
OECD. (2016a). Test No. 421: Reproduction/Developmental Toxicity Screening Test. OECD. https://doi.org/10.1787/9789264264380-en
OECD. (2016b). Test No. 422: Combined Repeated Dose Toxicity Study with the Reproduction/Developmental Toxicity Screening Test. OECD. https://doi.org/10.1787/9789264264403-en
OECD. (2018a). Test No. 414: Prenatal Developmental Toxicity Study. OECD. https://doi.org/10.1787/9789264070820-en
OECD. (2018b). Test No. 443: Extended One-Generation Reproductive Toxicity Study. OECD. https://doi.org/10.1787/9789264185371-en
OECD. (2023). Test No. 458: Stably Transfected Human Androgen Receptor Transcriptional Activation Assay for Detection of Androgenic Agonist and Antagonist Activity of Chemicals. OECD. https://doi.org/10.1787/9789264264366-en
Paulozzi, L. J. (1999). International trends in rates of hypospadias and cryptorchidism.
Robitaille, J., & Langlois, V. S. (2020). Consequences of steroid-5α-reductase deficiency and inhibition in vertebrates. General and Comparative Endocrinology, 290, 113400. https://doi.org/10.1016/j.ygcen.2020.113400
Sinclair, A., Cao, M., Pask, A., Baskin, L., & Cunha, G. (2017). Flutamide-induced hypospadias in rats: A critical assessment. Differentiation; Research in Biological Diversity, 94, 37–57. (rayyan-427634666). https://doi.org/10.1016/j.diff.2016.12.001
Skakkebaek, N. E., Rajpert-De Meyts, E., Louis, G. M. B., Toppari, J., Andersson, A.-M., Eisenberg, M. L., Jensen, T. K., Jorgensen, N., Swan, S. H., Sapra, K. J., Ziebe, S., Priskorn, L., & Juul, A. (2016). Male Reproductive Disorders And Fertility Trends: Influences Of Environement And Genetic susceptibility. PHYSIOLOGICAL REVIEWS, 96(1), 55–97. (WOS:000370383200003). https://doi.org/10.1152/physrev.00017.2015
Stewart, M. K., Mattiske, D. M., & Pask, A. J. (2018). In utero exposure to both high- and low-dose diethylstilbestrol disrupts mouse genital tubercle development†. Biology of Reproduction, 99(6), 1184–1193. https://doi.org/10.1093/biolre/ioy142
Svingen, T., Villeneuve, D. L., Knapen, D., Panagiotou, E. M., Draskau, M. K., Damdimopoulou, P., & O’Brien, J. M. (2021). A Pragmatic Approach to Adverse Outcome Pathway Development and Evaluation. Toxicological Sciences, 184(2), 183–190. https://doi.org/10.1093/toxsci/kfab113
Tut, T. G., Ghadessy, F. J., Trifiro, M. A., Pinsky, L., & Yong, E. L. (1997). Long Polyglutamine Tracts in the Androgen Receptor Are Associated with Reduced Trans -Activation, Impaired Sperm Production, and Male Infertility 1. The Journal of Clinical Endocrinology & Metabolism, 82(11), 3777–3782. https://doi.org/10.1210/jcem.82.11.4385
Welsh, M., Saunders, P. T. K., Fisken, M., Scott, H. M., Hutchison, G. R., Smith, L. B., & Sharpe, R. M. (2008). Identification in rats of a programming window for reproductive tract masculinization, disruption of which leads to hypospadias and cryptorchidism. Journal of Clinical Investigation, 118(4), 1479–1490. https://doi.org/10.1172/JCI34241
Williams, A. J., Grulke, C. M., Edwards, J., McEachran, A. D., Mansouri, K., Baker, N. C., Patlewicz, G., Shah, I., Wambaugh, J. F., Judson, R. S., & Richard, A. M. (2017). The CompTox Chemistry Dashboard: A community data resource for environmental chemistry. Journal of Cheminformatics, 9(1), 61. https://doi.org/10.1186/s13321-017-0247-6
Willingham, E., Agras, K., Souza, A. J. de, Konijeti, R., Yucel, S., Rickie, W., Cunha, G., & Baskin, L. (2006). Steroid receptors and mammalian penile development: An unexpected role for progesterone receptor? The Journal of Urology, 176(2), 728–733. (rayyan-427634662). https://doi.org/10.1016/j.juro.2006.03.078
Yucel, S., Liu, W., Cordero, D., Donjacour, A., Cunha, G., & Baskin, L. (2004). Anatomical studies of the fibroblast growth factor-10 mutant, Sonic Hedge Hog mutant and androgen receptor mutant mouse genital tubercle. Advances in Experimental Medicine and Biology, 545, 123–148. (rayyan-427634903). https://doi.org/10.1007/978-1-4419-8995-6_8
Zheng, Z., Armfield, B., & Cohn, M. (2015). Timing of androgen receptor disruption and estrogen exposure underlies a spectrum of congenital penile anomalies. Proceedings of the National Academy of Sciences of the United States of America, 112(52), E7194-203. (rayyan-427635801). https://doi.org/10.1073/pnas.1515981112