
This Key Event Relationship is licensed under the Creative Commons BY-SA license. This license allows reusers to distribute, remix, adapt, and build upon the material in any medium or format, so long as attribution is given to the creator. The license allows for commercial use. If you remix, adapt, or build upon the material, you must license the modified material under identical terms.
Relationship: 3486
Title
Decrease, circulating testosterone levels leads to nipple retention, increased
Upstream event
Downstream event
Key Event Relationship Overview
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding | Point of Contact | Author Status | OECD Status |
|---|---|---|---|---|---|---|
| Decreased testosterone synthesis leading to increased nipple retention (NR) in male (rodent) offspring | non-adjacent | Moderate | Terje Svingen (send email) | Under development: Not open for comment. Do not cite |
Taxonomic Applicability
Sex Applicability
| Sex | Evidence |
|---|---|
| Male | High |
Life Stage Applicability
| Term | Evidence |
|---|---|
| Foetal | High |
Key Event Relationship Description
This KER describes a fetal decrease in circulating testosterone (often measured in serum or plasma) leading to NR in male rodent offspring. In rats and mice, females develop 10 and 12 nipples, respectively, with males typically displaying zero. In male rodents, testosterone is primarily produced by the fetal testes, secreted into the bloodstream, and transported to the peripheral reproductive tissues, including the preliminary mammary tissue. Testosterone can bind directly to the AR in the tissue or first being converted to DHT by 5α-reductase (Murashima et al., 2015). AR activation by androgens in mesenchymal cells of the developing mammary glands causes cell death and subsequent separation of the tissue from the epidermis, resulting in no formation of nipples (Kratochwil, 1986). In females, where androgen levels are low, nipple formation is not blocked. The dependency of androgens for suppression of nipple development in males means that reductions in circulating testosterone levels can lead to retention of nipples.
In humans, both sexes have two nipples, and there is no known androgen-driven sexual dimorphism (Schwartz et al., 2021). The KER is thus not considered directly applicable to humans, but is a clear readout of reduced androgen action and fetal masculinization during development, which is relevant to humans (Schwartz et al., 2021). It is included as a mandatory endpoint in several rodent OECD Test Guidelines (OECD, 2025a, 2025b, 2025c) and considered an adverse outcome applicable to the setting of Points of Departure for use in human health risk assessment (OECD, 2013).
Evidence Collection Strategy
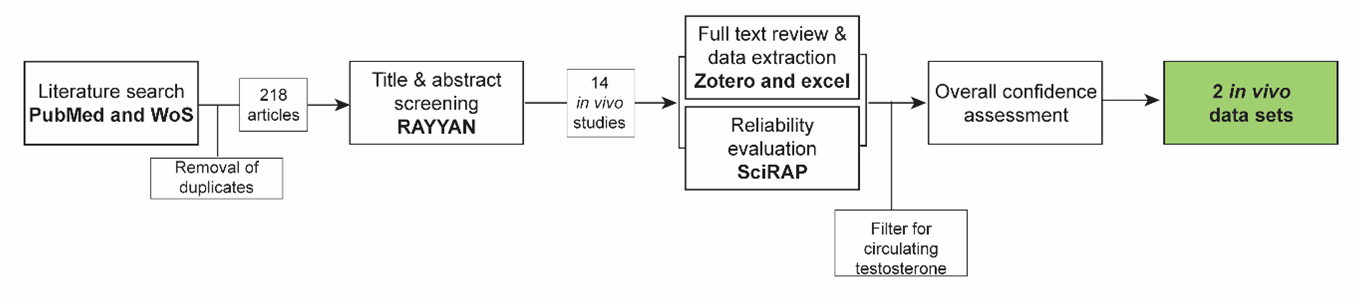
A systematic approach (Fig. 1, 570lkl0x3z_KER_3486_Figure_1.png (1355×301)) was used to collect evidence based on the methodology described in (Holmer et al., 2024). The evidence collection for this KER was done concurrently with the evidence collection for KER-3487 ‘Decreased, intratesticular testosterone leads to nipple retention, increased’, for which the same search string was used.
{kind=link}
Search strategy
Search strings were synthesized for PubMed and Web of Science based on the review question, ‘Does decreased testosterone during fetal development lead to decreased anogenital distance in male mammals?'
Search string in PubMed: "testosterone*" AND ("nipple*” OR “areola*”)
Search string in Web of Science: "testosterone*" AND ("nipple*” OR “areola*”)
The searches were performed on 01.10.2024
Title & abstract screening:
Retrieved articles were screened in the online tool RAYYAN https://www.rayyan.ai/. After the removal of duplicates, the titles, and abstracts of the remaining 218 articles were screened according to pre-defined inclusion and exclusion criteria:
Inclusion criteria:
- In vivo studies in male mammals where fetal testosterone is reduced and nipples / areolas are measured *
- Reviews on nipple/areola retention
- In vitro, ex vivo, and in vivo mechanistic studies on nipple/areola retention
Exclusion criteria:
- Papers not in English
- Abstracts and other non-full-text publications
*In cases where this criterion could not be determined by reading the abstract, the study was included for full text review.
Full text review, data extraction, and reliability evaluation of animal studies:
For the in vivo studies, the full-text papers were reviewed using the same exclusion criteria as in the title & abstract screening, and data were extracted from the included papers into an Excel template. In parallel, methodological reliability was assessed using the online tool Science in Risk Assessment and Policy (SciRAP; http://www.scirap.org, see appendix 1, 93nzlfrd7z_KER_3486_Appendix_1.pdf). Based on the SciRAP evaluations, animal studies were assigned a reliability category using the principles outlined in table 1. Studies were divided into different datasets if multiple different chemicals, different exposure windows, or different timepoints of measurement of NR were included.
Moreover, as this KER was made in parallel with several other KERs for other male reproductive endpoints (anogenital distance and hypospadias), five studies retrieved in the searches for these KERs, which also measured NR, but were not detected in the search for this KER were also added, data extracted and evaluated for reliability.
The collected data were then filtered to only include data sets measuring circulating testosterone, either in plasma or serum. Mixture studies were excluded from the final evidence as the chemicals may have many different upstream mechanisms.
Table 1 Principles for translation of SciRAP evaluations into reliability categories.
|
Reliability Category |
Principles for Categorization |
|
1.Reliable without restriction |
SciRAP methodological quality Score > 80 and all key criteriaa are “Fulfilled” and there are no deficiencies in the non-key criteria that might affect study reliability. |
|
2. Reliable with restriction |
SciRAP methodological quality Score > 65 and one or several of the key criteria are “Partially Fulfilled” or there are minor deficiencies in the non-key criteria that might affect study reliability. |
|
3. Not reliable |
SciRAP methodological quality Score < 65 or one or several of the key criteria are “Not Fulfilled” or there are major deficiencies in the non-key criteria that affect study reliability. |
|
4. Not assignable |
Two or more of the key criteria are “Not Determined” |
aKey criteria were SciRAP criteria for methodological quality, judged as specifically critical for reliability of the data for this KER and were determined a priori. The key criteria for this data collection are outlined in appendix 1 (93nzlfrd7z_KER_3486_Appendix_1.pdf).
Evidence Supporting this KER
Biological Plausibility
The biological plausibility for this KER is judged to be high, given the canonical biological knowledge on normal reproductive development in rodents.
Sexual differentiation in males, including blocking of nipple development in rodents, is programmed in fetal life. Once formed, the fetal testes synthesize testosterone through the steroidogenesis pathway. Testosterone is secreted and transported in the bloodstream either as free testosterone or bound to plasma proteins (albumin or sex-hormone binding globulin). Testosterone binds AR in peripheral tissues and can also be converted to DHT by the enzyme 5α-reductase. Binding of testosterone and DHT to AR program the fetal reproductive tissue to male differentiation (Murashima et al., 2015).
In most rodents, development of the nipples is a sexually dimorphic process. In female rats, where androgen levels are low, the nipples develop along the milk lines forming 12 nipples, which are visible around postnatal day 12-14 (Schwartz et al., 2021). In male rats, AR activation in fetal life suppresses the formation of the nipples through apoptosis of epithelial cells in the developing mammary glands. Normally, male rats do therefore not have nipples, although in rare occasions male control rats may display one or more retained nipples (Kratochwil, 1986; Schwartz et al., 2021).
Testosterone is produced from around GD15 in fetal rats and is present in circulation around the same time. Programming of the nipple tissue to regress mainly occurs within the masculinization programming window (GD16-20 in rats) (Welsh et al., 2014).
Given the dependency of testosterone for the regression of the nipples, either through direct AR activation or conversion to DHT, it is highly plausible that a decrease in circulating levels of testosterone will lead to nipple retention in males.
Empirical Evidence
The empirical evidence from studies in animals for this KER is overall judged as moderate
From the data collection, two data sets were extracted. The data set included two different stressors causing reduced fetal levels of circulating testosterone in rats (Table 2 and appendix 2, 61b9o238vs_KER_3486_Appendix_2.pdf). Both data sets showed concurrent NR.
Table 2 Empirical evidence for KER 3486 LOAEL: Lowest observed adverse effect level; See appendix 2 for specifications.
|
Species |
Stressors(s) |
Effect on upstream event (circulating testosterone) |
Effect on downstream event (NR) |
Reference |
|
Rat |
Diethylhexyl phthalate |
LOAEL 750 mg/kg bw/day |
LOAEL 750 mg/kg bw/day |
(Borch et al., 2004) |
|
Rat |
Prochloraz |
LOAEL 30 mg/kg bw/day |
LOAEL 30 mg/kg bw/day |
(Vinggaard et al., 2005) |
Dose concordance
The empirical evidence for this KER does not inform dose concordance, as no study uses more than one dose of stressor.
Temporal concordance
NR is generally first observable in postnatal animals, while the reductions in intratesticular testosterone are measured early in fetal life during exposure. This was demonstrated with prochloraz-induced NR, which was observed at PND13, but not when examining the males at GD21, when circulating testosterone levels were reduced (Vinggaard AM et al., 2005).
Incidence concordance
The data does not inform incidence concordance.
Uncertainties and Inconsistencies
The low number of studies retrieved in the empirical evidence collection, this KER is in itself an uncertainty, and both studies only investigated one stressor dose.
An uncertainty in the empirical evidence is that prochloraz is also known to be an AR antagonist (Andersen et al., 2002), and it can therefore not be excluded that the effects of prochloraz on NR is, at least partly, due to direct antagonism of AR and not due to the low testosterone levels.
Known modulating factors
| Modulating Factor (MF) | MF Specification | Effect(s) on the KER | Reference(s) |
|---|---|---|---|
| Rat strain | Long-Evans Hooded rats are less sensitive to NR than Sprague Dawley rats | (Wolf et al., 1999; You et al., 1998) |
Quantitative Understanding of the Linkage
The quantitative understanding of this KER is classified as low.
Response-response Relationship
There is no known information on the response-response relationship for this KER.
Time-scale
The time scale of this KER is weeks. Testosterone is secreted from ~GD15 in rats, which is at the beginning of the masculinization programming window. The mammary glands start developing during fetal life as well, but cannot be observed in female rodents until weeks after birth, which is also the time at which NR can be observed in males (Schwartz et al., 2021)
Known Feedforward/Feedback loops influencing this KER
There are no known feedback/feedforward loops for this KER.
Domain of Applicability
Taxonomic applicability
This KER is considered applicable to rodents (evidence primarily from laboratory rats and mice), where males normally lack nipples due to suppressed differentiation by high levels of androgens. The empirical evidence in this KER supports that reduction in testosterone causes NR in rats, while relevance in mice is assumed based on knowledge about developmental biology in this species. In humans, both sexes have two nipples, and there is no known androgen-driven sexual dimorphism (Schwartz et al., 2021). The KER is thus not considered directly applicable to humans. However, NR is a clear readout of reduced androgen action and fetal masculinization during development in rodents, which is relevant to humans (Schwartz et al., 2021). It is included as a mandatory endpoint in several rodent OECD Test Guidelines (OECD, 2025a, 2025b, 2025c) and considered an adverse outcome applicable to the setting of Points of Departure for use in human health risk assessment (OECD, 2013).
Sex applicability
This KER is only applicable to males, as female rats and mice develop 12 and 10 nipples, respectively (Schwartz et al., 2021). Females do have circulating testosterone in fetal life, but the levels are much lower than in males (Houtsmuller et al., 1995), and do therefore not suppress nipple formation.
Life stage applicability
The programming for androgen-driven suppression of nipple development in rodents occurs during fetal life, around gestational days (GD) 16-20 in rats (Imperato-McGinley et al., 1986). In both male and female rodents, the development of the mammary glands starts in fetal life, including initial growth and subsequent sexual differentiation (Kratochwil, 1986; Watson & Khaled, 2008). The relevant timing for the investigation of NR is PND12-14 in male rat offspring when the nipples are visible in the female littermates. At this time in development, the nipples/areolas are visible through the skin without excessive fur that may interfere with the investigation (Schwartz et al., 2021).
References
Andersen, H. R., Vinggaard, A. M., Rasmussen, T. H., Gjermandsen, I. M., & Bonefeld-Jørgensen, E. C. (2002). Effects of currently used pesticides in assays for estrogenicity, androgenicity, and aromatase activity in vitro. Toxicology and Applied Pharmacology, 179(1), 1–12. https://doi.org/10.1006/taap.2001.9347
Borch J, Ladefoged O, Hass U, & Vinggaard AM. (2004). Steroidogenesis in fetal male rats is reduced by DEHP and DINP, but endocrine effects of DEHP are not modulated by DEHA in fetal, prepubertal and adult male rats. Reproductive Toxicology (Elmsford, N.Y.), 18(1), 53–61. https://doi.org/10.1016/j.reprotox.2003.10.011
Holmer, M. L., Zilliacus, J., Draskau, M. K., Hlisníková, H., Beronius, A., & Svingen, T. (2024). Methodology for developing data-rich Key Event Relationships for Adverse Outcome Pathways exemplified by linking decreased androgen receptor activity with decreased anogenital distance. Reproductive Toxicology, 128, 108662. https://doi.org/10.1016/j.reprotox.2024.108662
Houtsmuller, E. J., de Jong, F. H., Rowland, D. L., & Slob, A. K. (1995). Plasma testosterone in fetal rats and their mothers on day 19 of gestation. Physiology & Behavior, 57(3), 495–499. https://doi.org/10.1016/0031-9384(94)00291-C
Imperato-McGinley J, Binienda Z, Gedney J, & Vaughan ED Jr. (1986). Nipple differentiation in fetal male rats treated with an inhibitor of the enzyme 5 alpha-reductase: definition of a selective role for dihydrotestosterone. Endocrinology, 118(1), 132–137. https://doi.org/10.1210/endo-118-1-132
Kratochwil, K. (1986). Tissue Combination and Organ Culture Studies in the Development of the Embryonic Mammary Gland. In R. B. L. Gwatkin (Ed.), Manipulation of Mammalian Development (pp. 315–333). Springer US. https://doi.org/10.1007/978-1-4613-2143-9_11
Murashima, A., Kishigami, S., Thomson, A., & Yamada, G. (2015). Androgens and mammalian male reproductive tract development. Biochimica et Biophysica Acta (BBA) - Gene Regulatory Mechanisms, 1849(2), 163–170. https://doi.org/10.1016/j.bbagrm.2014.05.020
OECD (2013), Guidance Document Supporting OECD Test Guideline 443 on the Extended One-Generational Reproductive Toxicity Test, OECD Series on Testing and Assessment, No. 151, OECD Publishing, Paris, ENV/JM/MONO(2013)10
OECD (2025a), Test No. 443: Extended One-Generation Reproductive Toxicity Study, OECD Guidelines for the Testing of Chemicals, Section 4, OECD Publishing, Paris, https://doi.org/10.1787/9789264185371-en.
OECD (2025b), Test No. 421: Reproduction/Developmental Toxicity Screening Test, OECD Guidelines for the Testing of Chemicals, Section 4, OECD Publishing, Paris, https://doi.org/10.1787/9789264264380-en.
OECD (2025c), Test No. 422: Combined Repeated Dose Toxicity Study with the Reproduction/Developmental Toxicity Screening Test, OECD Guidelines for the Testing of Chemicals, Section 4, OECD Publishing, Paris, https://doi.org/10.1787/9789264264403-en.
Schwartz CL, Christiansen S, Hass U, Ramhøj L, Axelstad M, Löbl NM, & Svingen T. (2021). On the Use and Interpretation of Areola/Nipple Retention as a Biomarker for Anti-androgenic Effects in Rat Toxicity Studies. Frontiers in Toxicology, 3, 730752. https://doi.org/10.3389/ftox.2021.730752
Vinggaard AM, Christiansen S, Laier P, Poulsen ME, Breinholt V, Jarfelt K, Jacobsen H, Dalgaard M, Nellemann C, & Hass U. (2005). Perinatal exposure to the fungicide prochloraz feminizes the male rat offspring. Toxicological Sciences : An Official Journal of the Society of Toxicology, 85(2), 886–897. https://doi.org/doi.org/10.1093/toxsci/kfi150
Watson, C. J., & Khaled, W. T. (2008). Mammary development in the embryo and adult: A journey of morphogenesis and commitment. Development, 135(6), 995–1003. https://doi.org/10.1242/dev.005439
Welsh M, Suzuki H, & Yamada G. (2014). The masculinization programming window. Endocrine Development, 27, 17–27. https://doi.org/10.1159/000363609
Wolf C Jr, Lambright C, Mann P, Price M, Cooper RL, Ostby J, & Gray LE Jr. (1999). Administration of potentially antiandrogenic pesticides (procymidone, linuron, iprodione, chlozolinate, p,p’-DDE, and ketoconazole) and toxic substances (dibutyl- and diethylhexyl phthalate, PCB 169, and ethane dimethane sulphonate) during sexual differentiation produces diverse profiles of reproductive malformations in the male rat. Toxicology and Industrial Health, 15(1), 94–118. https://doi.org/10.1177/074823379901500109
You L, Casanova M, Archibeque-Engle S, Sar M, Fan LQ, & Heck HA. (1998). Impaired male sexual development in perinatal Sprague-Dawley and Long-Evans hooded rats exposed in utero and lactationally to p,p’-DDE. Toxicological Sciences : An Official Journal of the Society of Toxicology, 45(2), 162–173. https://doi.org/10.1093/toxsci/45.2.162