
This Key Event Relationship is licensed under the Creative Commons BY-SA license. This license allows reusers to distribute, remix, adapt, and build upon the material in any medium or format, so long as attribution is given to the creator. The license allows for commercial use. If you remix, adapt, or build upon the material, you must license the modified material under identical terms.
Relationship: 3717
Title
Increased, E2 production in ovaries leads to Plasma E2, increased
Upstream event
Downstream event
Key Event Relationship Overview
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding | Point of Contact | Author Status | OECD Status |
|---|---|---|---|---|---|---|
| Activation, estrogen receptor alpha leads to persistent vaginal cornification via increased kisspeptin release | adjacent | High | John Frisch (send email) | Under development: Not open for comment. Do not cite | ||
| Activation, estrogen receptor alpha leads to increased uterine weight via earlier proliferation of cells of the uterine lining | adjacent | High | John Frisch (send email) | Under development: Not open for comment. Do not cite | ||
| Activation, estrogen receptor alpha leads to precocious puberty via increased kisspeptin release | adjacent | High | John Frisch (send email) | Under development: Not open for comment. Do not cite | ||
| Activation, estrogen receptor alpha leads to decreased fertility via prolonged estrus cycle | adjacent | High | John Frisch (send email) | Under development: Not open for comment. Do not cite |
Taxonomic Applicability
| Term | Scientific Term | Evidence | Link |
|---|---|---|---|
| mammals | mammals | Moderate | NCBI |
Sex Applicability
| Sex | Evidence |
|---|---|
| Female | High |
Life Stage Applicability
| Term | Evidence |
|---|---|
| Adult, reproductively mature | Moderate |
| Juvenile | Moderate |
Key Event Relationship Description
Production of estradiol (E2) by the ovaries has been well-established by the two-cell, two gonadotropin model of steroid biosynthesis (for review see Drummond 2006; Kimura et al. 2007; Palermo 2007; Beevors et al. 2024). Luteinizing hormone stimulates steroid production in theca cells, with follicle-stimulating hormone stimulates steroid production in granulosa cells.
Estradiol is a key signalling hormone, with increased production of estradiol in the ovary leading to increased secretion of estradiol into plasma.
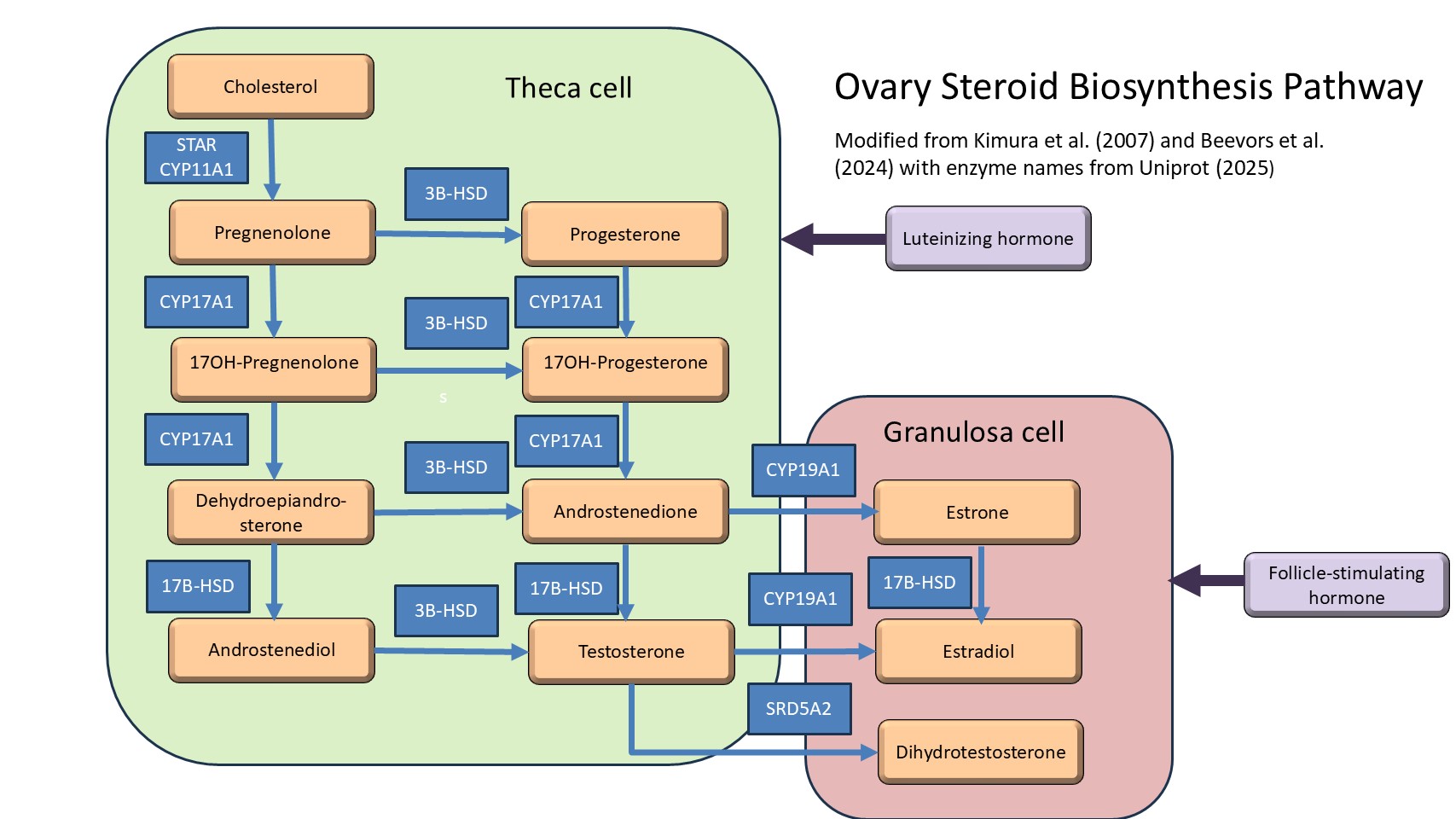
Table 1: List of steroid synthesis enzymes with identifier of enzyme (Uniprot, 2025).
|
Enzyme |
Identifier |
|
Steroidogenic acute regulatory protein, mitochondrial (STAR) |
|
|
Cholesterol side-chain cleavage enzyme, mitochondrial (CYP11A) |
EC:1.14.15.6 |
|
3 beta-hydroxysteroid dehydrogenase (3B-HSD) |
EC:1.1.1.145 |
|
Steroid 17-alpha-hydroxylase (CYP17A1) |
EC:1.14.14.19 |
|
17-beta-hydroxysteroid dehydrogenase (17B-HSD) |
EC:1.1.1.105 |
|
Aromatase (CYP19A1) |
EC:1.14.14.14 |
|
3-oxo-5-alpha-steroid 4-dehydrogenase 2 (SRD5A2) |
EC:1.3.1.22 |
Evidence Collection Strategy
This Key Event Relationship was part of an Environmental Protection Agency effort to develop AOPs that establish scientifically supported causal linkages between alternative endpoints measured using new approach methodologies (NAMs) and guideline apical endpoints measured in Tier 1 and Tier 2 test guidelines (U.S. EPA, 2025) employed by the Endocrine Disruptor Screening Program (EDSP). A series of key events that represent significant, measurable, milestones connecting molecular initiation to apical endpoints indicative of adversity were identified based on scientific review articles and empirical studies. Additionally, scientific evidence supporting the causal relationships between each pair of key events was assembled and evaluated. The present effort focused primarily on empirical studies with laboratory rodents and other mammals.
Empirical studies are focused on increased estradiol production in ovaries and resulting increased plasma estradiol, in support of development of AOP 623.
Authors of KER 3717 did a further evaluation of published peer-reviewed literature to provide additional evidence in support of the key event relationship. The literature used to support this KER began with the test guidelines and followed to primary, secondary, and/or tertiary works concerning the relevant underlying biology. In addition, search engines were used to target journal articles with term ‘Estradiol’ to locate representative empirical studies that support the key event relationship.
Evidence Supporting this KER
Biological Plausibility
Increased estradiol production in gonads and resulting increased plasma estradiol have been studied in laboratory mammals by addition of hormones (Sashinda and Johnson 1976) and toxicants with endocrine disrupting properties (Li et al. 2018; Gan et al. 2024). Studies that link increases in steroidogenic enzymes in ovaries to increased estradiol levels in plasma have been useful in establishing plausibility (Li et al. 2018; Gan et al. 2024). Ovaries have been established as the primary source of estradiol in females by the two-cell, two gonadotropin model of steroid biosynthesis (Drummond 2006; Kimura et al. 2007; Palermo 2007; Beevors et al. 2024).
Empirical Evidence
|
Species |
Duration |
Dose |
Increased E2 production ovaries? |
Increased plasma estradiol? |
Summary |
Citation |
|
Rats (Rattus norvegicus) |
24 hours |
10 ug/L FSH and 1 ug/L LH at 2 hour intervals. |
yes |
yes |
Female rats injected with FSH and LH had increased estradiol production in ovaries leading to increased plasma estradiol. |
Sashinda and Johnson (1976) |
|
Rats (Rattus norvegicus) |
3 generations |
1, 5 mg/kg BW Cadmium chloride |
yes |
yes |
Female rats exposed to cadmium chloride had increased estradiol production in ovaries and statistically significant increased levels of steroidogenic enzymes (STAR, CYP11A1, 3B-HSD, CYP19A1) primarily at 5 mg/kg BW leading to statistically significant increased plasma estradiol at 5 mg/kg BW. |
Li et al. (2018) |
|
Mice (Mus musculus) |
Until post natal day 55 |
30, 300, 3000 ug/kg/d triclosan |
yes |
yes |
Female mice exposed to triclosan increased estradiol production in ovaries by increased statistically significant expression of steroidogenic enzyme genes (STAR at >= 300 ug/kg/d, CYP11A1 at >= 30 ug/kg/d, CYP17A1 at 3000 ug/kg/d, CYP19A1 at >= 300 ug/kg/d) leading to statistically significant increased plasma estradiol at >= 300 ug/kg/d. |
Gan et al. (2024) |
Uncertainties and Inconsistencies
Known modulating factors
Quantitative Understanding of the Linkage
Response-response Relationship
Time-scale
Known Feedforward/Feedback loops influencing this KER
Domain of Applicability
Life Stage: Applies to adult, reproductively mature and juveniles.
Sex: Applies to females as specific to ovaries.
Taxonomic: Primarily studied in humans and laboratory rodents. Plausible for most mammals due to conserved hormone pathways regulating hypothalamus-pituitary-gonadal axis processes. Estradiol production in ovaries widespread among vertebrates, including mammals (Bondesson et al. 2015), birds (Hanlon et al. 2022), fish (Li et al. 2019), reptiles (Cruz-Cano et al. 2023), and amphibians (Bondesson et al. 2015), with corresponding increases in plasma estradiol resulting from release of estradiol from ovaries.
References
Beevors LI, Sundar S, Foster PA. 2024. Steroid metabolism and hormonal dynamics in normal and malignant ovaries. Essays in Biochemistry 68(4): 491-507.
Bondesson M, Hao R, Lin CY, Williams C, Gustafsson JA. 2015. Estrogen receptor signaling during vertebrate development. Biochimica et Biophysica Acta 1849(2): 142-151.
Cruz-Cano NB, Sanchez-Rivera UA, Alvarez-Rodriguez C, Cardenas-Leon M, Martinez-Torres M. 2023. Sex steroid receptors in the ovarian follicles of the lizard Sceloporus torquatus. Zygote. 31(4): 386-392.
Drummond AE. 2006. The role of steroids in follicular growth. Reproductive Biology and Endocrinology 4:16.
Gan H, Lan H, Hu Z, Zhu B, Sun L, Jiang Y, Wu L, Liu J, Ding Z, Ye X. 2024. Triclosan induces earlier puberty onset in female mice via interfering with L-type calcium channels and activating Pik3cd. Ecotoxicology and Environmental Safety 269: 115772.
Hanlon C, Ziezold CJ, Bedecarrats GY. 2022. The Diverse Roles of 17β-Estradiol in Non-Gonadal Tissues and Its Consequential Impact on Reproduction in Laying and Broiler Breeder Hens. Frontiers in Physiology 13: 942790.
Kimura S, Matsumoto T, Matsuyama R, Shiina H, Sato T, Takeyama K, Kato S. 2007. Androgen receptor function in folliculogenesis and its clinical implication in premature ovarian failure. Trends in Endocrinology and Metabolism 18(5): 183-189.
Li M, Sun L, Wang D. 2019. Roles of estrogens in fish sexual plasticity and sex differentiation. General and Comparative Endocrinology 277: 9-16.
Li Z, Li T, Leng Y, Chen S, Liu Q, Feng J, Chen H, Huang Y, Zhang Q. 2018. Hormonal changes and folliculogenesis in female offspring of rats exposed to cadmium during gestation and lactation. Environmental Pollution 238: 336-347.
Palermo R. 2007. Differential actions of FSH and LH during folliculogenesis. Reproductive BioMedicine Online 15(3): 326-337.
Sashida T, Johnson DC. 1976. Stimulation of the estrogen synthesizing system of the immature rat ovary by exogenous and endogenous gonadotropins. Steroids 27(4): 469-79.
U.S. Environmental Protection Agency. 2025. EDSP Test Guidelines and Guidance Document. https://www.epa.gov/test-guidelines-pesticides-and-toxic-substances/edsp-test-guidelines-and-guidance-document (retrieved 25 July 2025).
The UniProt Consortium. UniProt: the Universal Protein Knowledgebase in 2025. https://www.uniprot.org/ (retrieved 2 November 2025).
Italics indicate edits from John Frisch February 2026. A full list of updates can be found in the Change Log on the View History page.