
This AOP is licensed under the BY-SA license. This license allows reusers to distribute, remix, adapt, and build upon the material in any medium or format, so long as attribution is given to the creator. The license allows for commercial use. If you remix, adapt, or build upon the material, you must license the modified material under identical terms.
AOP: 576
Title
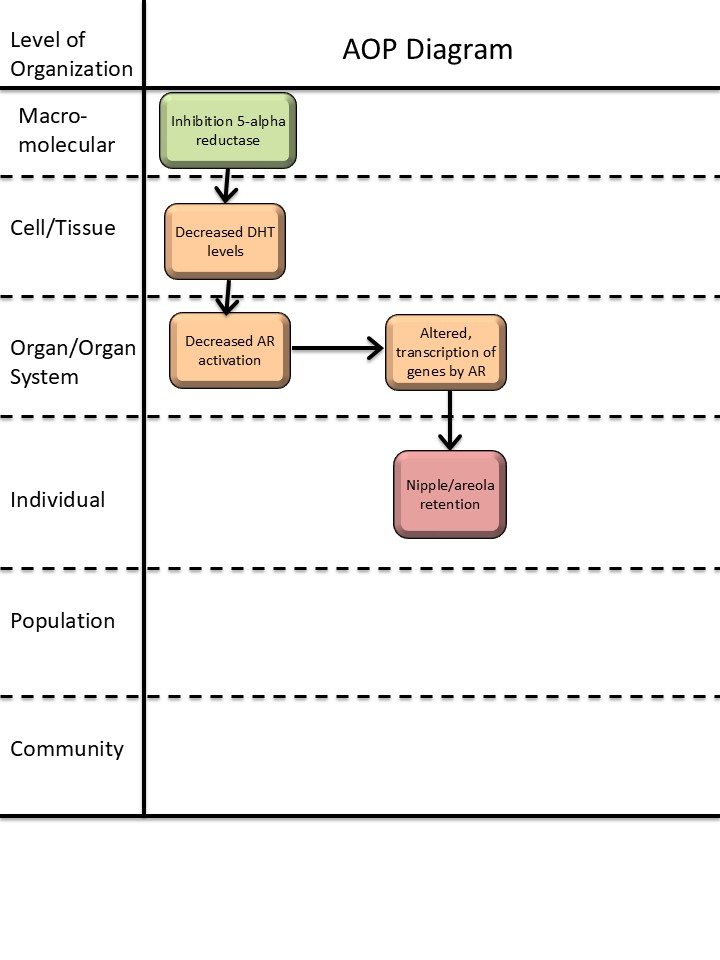
5α-reductase inhibition leading to increased nipple retention (NR) in male (rodent) offspring
Short name
Graphical Representation
Additional AOP Exploration Options
Click links below to explore AOP 576, 5α-reductase inhibition leading to increased nipple retention (NR) in male (rodent) offspring in tools offered by third parties.
Point of Contact
Contributors
- Emilie Elmelund
- Terje Svingen
Coaches
OECD Information Table
| OECD Project # | OECD Status | Reviewer's Reports | Journal-format Article | OECD iLibrary Published Version |
|---|---|---|---|---|
| Scientific Review |
This AOP was last modified on March 20, 2026 10:55
Revision dates for related pages
| Page | Revision Date/Time |
|---|---|
| Inhibition, 5α-reductase | March 20, 2026 11:45 |
| Decrease, dihydrotestosterone (DHT) levels | March 20, 2026 11:52 |
| Decrease, androgen receptor activation | March 20, 2026 11:54 |
| Altered, Transcription of genes by the androgen receptor | March 20, 2026 12:02 |
| Nipple retention (NR), increased | March 21, 2026 09:18 |
| Inhibition, 5α-reductase leads to Decrease, DHT level | March 20, 2026 12:20 |
| Decrease, AR activation leads to nipple retention, increased | March 20, 2026 11:08 |
| Decrease, DHT level leads to Decrease, AR activation | March 20, 2026 12:41 |
| Decrease, AR activation leads to Altered, Transcription of genes by the AR | March 20, 2026 10:30 |
| Finasteride | November 29, 2016 18:42 |
Abstract
This AOP links inhibition of 5α-reductase during fetal life with increased nipple/areola retention (NR) in male rodent offspring. NR, measured around 2 weeks postpartum, is a marker for disrupted masculinization of male rodents (primarily investigated in laboratory rats and mice) and is associated with general feminization of male offspring.
5α-reductase is an enzyme that converts testosterone to dihydrotestosterone (DHT). In normal male reproductive development, DHT activates the androgen receptor (AR) in many peripheral reproductive tissues to drive differentiation of the male phenotype, including regression of nipple anlagen in male rats and mice. While testosterone also acts directly at the AR, DHT is 5-10 times more potent and in tissues peripheral to the testes, conversion to DHT is necessary for proper masculinization (Amato et al., 2022; Davey & Grossmann, 2016).
This AOP delineates the evidence that inhibition of 5α-reductase reduces DHT levels and consequently AR activation, causing retention of nipples in male rodents. The AOP is supported by in vitro experiments upstream of AR activation and by in vivo studies downstream of AR activation. Downstream of a reduction in AR activation, the molecular mechanisms of NR are unclear, highlighting a knowledge gap in this AOP and potential for further development.
The confidence in each of the KERs comprising the AOP is judged as high, with both high biological plausibility and high confidence in empirical evidence. The mechanistic link between KE-286 (‘altered, transcription of genes by AR’) and AO 1786 (‘increase, nipple retention’) is not established, but given the high confidence in the KERs, the overall confidence in the AOP is judged as high.
The AOP supports the regulatory application of NR as a measure of endocrine disruption relevant for human health and the use of NR as an indicator of anti-androgenicity in environmentally relevant species. Even though NR cannot be directly translated to a human endpoint, the AOP is considered human relevant since NR is a clear readout of reduced androgen action and masculinization during development and is considered an ‘adverse outcome’ in OECD test guidelines (TG 443, TG 421, TG 422). The AOP also holds utility for informing on anti-androgenicity more generally, as this modality is highly relevant across mammalian species and vertebrates more broadly due to the conserved nature of the AR and its implication in sexual differentiation across species.
AOP Development Strategy
Context
This AOP is a part of an AOP network for reduced AR activation leading to increased NR in male offspring. The other AOPs in this network are AOP 344 (‘Androgen receptor antagonism leading to increased nipple retention in male (rodent) offspring’), and AOP 575 (‘Decreased testosterone synthesis leading to increased nipple retention in male (rodent) offspring’). The purpose of the AOP network is to organize the well-established evidence for anti-androgenic mechanisms-of-action leading to increased NR. It can be used in identification and assessment of endocrine disruptors and to inform predictive toxicology, identification of knowledge gaps for investigation and method development.
This work received funding from the European Food and Safety Authority (EFSA) under Grant agreement no. GP/EFSA/PREV/2022/01.
Strategy
The OECD AOP Developer’s Handbook v2.7 was followed alongside pragmatic approaches (Svingen et al., 2021).
The adverse outcome, AO 1786: Increase, nipple retention, was originally developed in (Pedersen et al., 2022).
Upstream KEs and KERs were developed in Draskau et al., 2024 as part of an anti-androgenic network, using mainly key review publications since it was considered canonical knowledge. This included MIE 1617: Inhibition, 5α-reductase; KE 1613: Decrease, DHT levels; KE 1614: Decrease, AR activation and KE 286: Altered transcription of genes by the AR as well as the connecting KERs, KER 1880: Inhibition, 5α-reductase leads to decrease, DHT levels; KER 1935: Decrease, DHT levels leads to decrease, AR activation and KER 2124: Decrease, AR activation leads to altered, transcription of genes by AR (Draskau et al., 2024).
The non-adjacent KER3348 linking reduced AR activation with NR was developed using a systematic weight-of-evidence approach, following the methodology outlined in (Holmer et al., 2024). Publications were retrieved by literature searches in PubMed and Web of Science and extensive screening using pre-defined inclusion and exclusion criteria. Evaluation of methodological reliability of in vivo animal studies was performed using the Science in Risk Assessment and Policy (SciRAP) online tool. As there are currently no in vivo methods to measure AR activation in mammals, six chemicals with known anti-androgenic mechanisms-of-action were chosen for the empirical evidence for this KER.
The rationale for the inclusion of KEs and KERs in the upstream anti-androgenic network is detailed in (Draskau et al., 2024). The link between the upstream network, more specifically KE 286 (‘altered, transcription of genes by AR’), and AO1786 (‘increase, nipple retention’) likely contains a tissue-specific KE that has not been developed, as sufficient evidence is not yet available. Thus, for now, the most evidence for the link between the upstream anti-androgenic network and increased NR is captured by KER 3348.
Summary of the AOP
Events:
Molecular Initiating Events (MIE)
Key Events (KE)
Adverse Outcomes (AO)
| Type | Event ID | Title | Short name |
|---|
| MIE | 1617 | Inhibition, 5α-reductase | Inhibition, 5α-reductase |
| KE | 1613 | Decrease, dihydrotestosterone (DHT) levels | Decrease, DHT level |
| KE | 1614 | Decrease, androgen receptor activation | Decrease, AR activation |
| KE | 286 | Altered, Transcription of genes by the androgen receptor | Altered, Transcription of genes by the AR |
| AO | 1786 | Nipple retention (NR), increased | nipple retention, increased |
Relationships Between Two Key Events (Including MIEs and AOs)
| Title | Adjacency | Evidence | Quantitative Understanding |
|---|
| Decrease, AR activation leads to nipple retention, increased | non-adjacent | High |
Network View
Prototypical Stressors
| Name |
|---|
| Finasteride |
Life Stage Applicability
| Life stage | Evidence |
|---|---|
| Foetal | High |
Taxonomic Applicability
Sex Applicability
| Sex | Evidence |
|---|---|
| Male | High |
Overall Assessment of the AOP
Domain of Applicability
The upstream part of the AOP has a broad applicability domain, but KER 3348 (Decrease, AR activation, leads to increased nipple retention) is considered only directly applicable to male rodent (current evidence primarily from rats and mice) during fetal life, restricting the applicability of the AOP. NR is specific to animals with sexual dimorphism in the number of nipples, a feature most prominently investigated in laboratory rats and mice. It is, however, biologically plausible that the AOP is applicable to other rodent species. The process of retention of nipples by disruption of androgen programming happens in the fetal life stage, but the AO is detected postnatally. In the males of mice and rats, the nipple anlagen are programmed during fetal development by androgens to regress, leading to no visible nipples in males postnatally, while females exhibit nipples. Specifically, the MPW (~gestational days (GD) 16-20 in rat, presumably gestational weeks (GW) 8-14 in humans) is the primary fetal window of applicability, but effects outside of this window in fetal life, after androgen production has started, cannot be excluded. This AOP only contains empirical evidence for the applicability to male rats, but the AOP is considered equally applicable to male mice, as these also normally exhibit nipple regression stimulated by androgens. Moreover, the AOP is relevant for other taxa, including humans, as NR in male rodents indicates a reduction in fetal masculinization. NR is therefore included as a mandatory endpoint in multiple OECD Test Guideline studies for developmental and reproductive toxicity and is considered applicable as an adverse outcome to set NOAELs and LOAELs of substances in human health risk assessments.
Essentiality of the Key Events
|
Event |
Evidence |
Uncertainties, inconsistencies and contradictory evidence |
|
MIE-1617 Inhibition, 5α-reductase HIGH: This MIE is usually measured in vitro, whereas the downstream events in the AOP are usually measured in vivo. Canonical knowledge of normal male reproductive development provides strong support for essentiality, along with 5α-reductase knockout models and models using exposure to 5α-reductase inhibitors. |
Biological plausibility provides strong support for the essentiality of this event, as DHT, produced by 5α-reductase, is a ligand of the AR and a primary driver of normal regression of nipple anlagen in male fetuses (Imperato-McGinley et al., 1986). Indirect evidence of impact of inhibition of 5α-reductase (MIE-1617) in vitro on AR activity in vitro: • Finasteride, a specific inhibitor of 5α-reductase, can decrease proliferation of prostate cancer cells in vitro, a proxy read-out of AR activity (Bologna et al., 1995). Direct evidence of impact of inhibition of 5α-reductase (MIE-1617) in vivo on decreased DHT levels (KE-1613): • Lack of 5α-reductase type 2 activity by e.g. inhibitor or KO decrease DHT levels locally in tissues and blood. This is demonstrated in humans, rats, monkeys, and mice (Robitaille & Langlois, 2020). Indirect evidence of impact of inhibition of 5α-reductase (MIE-1617) in vivo on decreased DHT levels (KE-1613): • Men with androgenic alopecia treated with finasteride or dutasteride presented with decreased DHT levels in serum (Clark et al., 2004; Drake et al., 1999). Direct evidence of impact of inhibition of 5α-reductase (MIE-1617) in vivo on increased nipple retention (AO-1786): • Exposure to the 5α-reductase inhibitors leads to increased retention of nipples in male offspring after in utero exposure (Christiansen et al., 2009; Imperato-McGinley et al., 1986). |
|
|
KE-1613 Decreased, DHT levels HIGH: Canonical knowledge of normal male reproductive development provides strong support for essentiality, along with rescue studies specifically demonstrating how DHT is essential for normal regression of nipple anlagen in male offspring.
|
Biological plausibility provides strong support for the essentiality of this event. Androgens are AR ligands and main drivers for the regression of nipple anlagen in male offspring (Goldman et al., 1976), with DHT playing an important role (Imperato-McGinley et al., 1986). Indirect evidence of impact of decreased DHT levels (KE-1613) on AR activation in vivo (KE-1614): • Androgen deprivation is used as treatment for prostate cancer, including 5α-reductase inhibitors, to reduce DHT levels and cancer growth (Aggarwal et al., 2010). Indirect evidence of impact of decreased DHT levels (KE-1613) on AR activation in vitro: • Increasing concentrations of DHT lead to increasing AR activation in vitro in AR reporter gene assays (OECD, 2023; Williams et al., 2017). Indirect evidence of impact of decreased DHT levels (KE-1613) on AR activity in vivo: • 5α-reductase 2 deficiency is an autosomal recessive condition in which 46,XY subjects with bilateral testes and normal testosterone production have impaired virilization during fetal life due to diminished DHT (Mendonca et al., 2016). Direct evidence of impact of decreased DHT levels (KE-1613) on increased nipple retention (AO-1786). • Nipple formation is inhibited in female rat fetuses exposed to DHT during gestation (Goldman et al., 1976). • Exposure to the 5α-reductase inhibitor 390 MSD leads to increased retention of nipples in male rats after in utero exposure, whereas simultaneous exposure to DHT reverses the effects (Imperato-McGinley et al., 1986). |
|
|
KE-1614 Decreased, AR activation HIGH: There is experimental evidence from mutant mice insensitive to androgens showing that the AR is essential for nipple retention in male offspring. There is also evidence from exposure studies in animals that substances antagonizing AR induce nipple retention in male pups. |
Biological plausibility provides strong support for the essentiality of this event, as AR activation is critical for normal regression of nipple anlagen in male embryos. Indirect evidence of the impact of decreased AR activation (KE-1614) on altered gene transcription by AR (KE-286): • Exposure to known anti-androgenic chemicals induces a changed gene expression pattern, e.g. in neonatal pig ovaries (Knapczyk-Stwora et al., 2019). Direct evidence of the impact of decreased AR activation (KE-1614) on altered gene transcription by AR (KE-286): • Male AR KO mice have altered gene expression pattern in a broad range of organs (see KER-2124). Indirect evidence of impact of decreased AR activation (KE-1614) on increased nipple retention (AO-1786): • Rat in vivo exposure to vinclozolin, procymidone and flutamide, which are known AR antagonists, leads to increased nipple retention in offspring (see KER-3348). Direct evidence of impact of decreased AR activation (KE-1614) on increased nipple retention (AO-1786): • Male Tfm mutant mice, which are insensitive to androgens and believed to be so due to a nonfunctional androgen receptor, present with retained nipples (Kratochwil & Schwartz, 1976) |
|
|
KE-286 Altered, trans. of genes by AR LOW: Strongest support for essentiality comes from biological plausibility. However, exact transcriptional effects and causality remain to be fully characterized. |
Biological plausibility provides support for the essentiality of this event. AR is a nuclear receptor and transcription factor regulating transcription of genes, and androgens, acting through AR, are essential for normal regression of nipple anlagen in male fetuses. |
There are currently no AR-responsive genes proved to be causally involved in nipple retention, and it is known that AR can also signal through non-genomic actions (Leung & Sadar, 2017). |
|
Event |
Direct evidence |
Indirect evidence |
Contradictory evidence |
Overall essentiality assessment |
|
MIE-1617 |
*** |
** |
|
High |
|
KE-1613 |
*** |
** |
|
High |
|
KE-1614 |
*** |
*** |
|
High |
|
KE-286 |
|
|
|
Low (biological plausibility) |
*Low level of evidence (some support for essentiality), ** Intermediate level of evidence (evidence for impact on one or more downstream KEs), ***High level of evidence (evidence for impact on AO).
Evidence Assessment
The confidence in each of the KERs comprising the AOP is judged as high, with both high biological plausibility and high confidence in empirical evidence. The mechanistic link between KE-286 (‘altered, transcription of genes by AR’) and AO 1786 (‘increase, nipple retention’) is not established, but given the high confidence in the KERs, the overall confidence in the AOP is judged as high.
|
KER |
Biological Plausibility |
Empirical Evidence |
Rationale |
|
KER-1880 Inhibition, 5α-reductase leads to a decrease, DHT levels |
High |
High (canonical) |
It is well established that 5α-reductase converts testosterone to DHT. In vitro, in vivo and human studies with 5α-reductase inhibitors have shown that the stressors dose-dependently decrease formation of DHT. |
|
KER-1935 Decrease, DHT levels leads to a decrease, AR activation |
High |
High (canonical) |
It is well established that DHT activates the AR. Direct evidence for this KER is not possible since KE 1614 can currently not be measured and is considered an in vivo effect. Indirect evidence using proxy read-outs of AR activation, either in vitro or in vivo, strongly supports the relationship. |
|
KER-2124 Decrease, AR activation leads to altered transcription of genes by AR |
High |
High (canonical) |
It is well established that the AR regulates gene transcription. In vivo animal studies and human genomic profiling show tissue-specific changes to gene expression upon disruption of AR. |
|
KER-3348 Decrease, AR activation leads to increase, nipple retention |
High |
High |
It is well established that activation of AR drives regression of nipple anlagen in males. The empirical evidence includes numerous in vivo toxicity studies showing that decreased AR activation leads to increased NR in male offspring, with few inconsistencies. The empirical evidence combined with theoretical considerations provide some support for dose, temporal, and incidence concordance for the KER, although this evidence is weak and indirect. It should be recognized that the upstream KE-1614 cannot currently be measured directly (in vivo). Instead, empirical evidence was therefore collected for substances known to affect upstream events. This limitation is not considered to lower the strength of the evidence in this case. |
Known Modulating Factors
|
Modulating factor (MF) |
Influence or Outcome |
KER(s) involved |
|
Genotype |
Extended CAG repeat length in AR is associated with reduced AR activity (Chamberlain et al., 1994; Tut et al., 1997). This MF could initiate the AOP at lower stressor doses. |
KER-1935, KER-2124, KER-3348 |
|
Rat strain |
Long-Evans Hooded rat is less sensitive to NR than the Sprague-Dawley rat (Wolf et al., 1999; You et al., 1998). Thus, the effects on AO at certain stressor doses may vary between strains |
KER-3348 |
Quantitative Understanding
The quantitative understanding of the AOP is limited. A key difficulty lies in the challenge of extrapolating from in vitro to in vivo events since these cannot be captured within the same experimental framework. Specifically, MIE-1617 is evaluated in vitro, while both KE-1613 (decrease, DHT levels’), KE-1614 (decrease, AR activation’) and the AO (Increase, NR) are in vivo endpoints. It should be noted that KE-1614 pertains to AR activation in vivo - currently lacking viable methods for direct measurement.
For in vivo to in vivo KERs like KER-1935 (‘Decrease, DHT level leads to Decrease, AR activation’) and KER-2124 (‘Decrease, AR activation leads to Altered, Transcription of genes by the AR’), there is not enough data to define a quantitative relationship, and such a relationship will differ between biological systems (species, tissue, cell type, life stage etc).
Considerations for Potential Applications of the AOP (optional)
The AOP supports the regulatory application of NR as a measure of endocrine disruption relevant for human health and the use of NR as an indicator of anti-androgenicity in mammals and other vertebrates in the environment. NR is a mandatory endpoint in multiple OECD test guidelines, including TG 443 (extended one-generation reproductive toxicity study) and TGs 421/422 (reproductive toxicity screening studies) (OECD 2025a; OECD 2025b; OECD 2025c). NR can contribute to establishing a No Observed Adverse Effect Level (NOAEL), as outlined in OECD guidance documents No. 43 and 151 (OECD 2008; OECD 2013). The ability to derive a NOAEL for increased NR in male rodent offspring, which can serve as a point of departure for determining human safety thresholds, underscores the regulatory significance of this AOP. The AOP also holds utility for informing on anti-androgenicity more generally, as this modality is highly relevant across mammalian species (Schwartz et al., 2021) and vertebrates more broadly due to the conserved nature of the AR and its implication in sexual differentiation across species (Ogino et al., 2023).
References
Aggarwal, S., Thareja, S., Verma, A., Bhardwaj, T. R., & Kumar, M. (2010). An overview on 5α-reductase inhibitors. Steroids, 75(2), 109–153. https://doi.org/10.1016/j.steroids.2009.10.005
Amato, C. M., Yao, H. H.-C., & Zhao, F. (2022). One Tool for Many Jobs: Divergent and Conserved Actions of Androgen Signaling in Male Internal Reproductive Tract and External Genitalia. Frontiers in Endocrinology, 13. https://doi.org/10.3389/fendo.2022.910964
Bologna, M., Muzi, P., Biordi, L., Festuccia, C., & Vicentini, C. (1995). Finasteride dose-dependently reduces the proliferation rate of the LnCap human prostatic cancer cell line in vitro. Urology, 45(2), 282–290. https://doi.org/10.1016/0090-4295(95)80019-0
Chamberlain, N. L., Driver, E. D., & Miesfeld, R. L. (1994). The length and location of CAG trinucleotide repeats in the androgen receptor N-terminal domain affect transactivation function. Nucleic Acids Research, 22(15), 3181–3186. https://doi.org/10.1093/nar/22.15.3181
Christiansen S, Scholze M, Dalgaard M, Vinggaard AM, Axelstad M, Kortenkamp A, & Hass U. (2009). Synergistic disruption of external male sex organ development by a mixture of four antiandrogens. Environmental Health Perspectives, 117(12), 1839–1846. https://doi.org/10.1289/ehp.0900689
Clark, R. V., Hermann, D. J., Cunningham, G. R., Wilson, T. H., Morrill, B. B., & Hobbs, S. (2004). Marked Suppression of Dihydrotestosterone in Men with Benign Prostatic Hyperplasia by Dutasteride, a Dual 5α-Reductase Inhibitor. The Journal of Clinical Endocrinology & Metabolism, 89(5), 2179–2184. https://doi.org/10.1210/jc.2003-030330
Davey, R. A., & Grossmann, M. (2016). Androgen Receptor Structure, Function and Biology: From Bench to Bedside. The Clinical Biochemist. Reviews, 36(4), 3–15.
Drake, L., Hordinsky, M., Fiedler, V., Swinehart, J., Unger, W. P., Cotterill, P. C., Thiboutot, D. M., Lowe, N., Jacobson, C., Whiting, D., Stieglitz, S., Kraus, S. J., Griffin, E. I., Weiss, D., Carrington, P., Gencheff, C., Cole, G. W., Pariser, D. M., Epstein, E. S., … Waldstreicher, J. (1999). The effects of finasteride on scalp skin and serum androgen levels in men with androgenetic alopecia. Journal of the American Academy of Dermatology, 41(4), 550–554.
Draskau, M. K., Rosenmai, A. K., Bouftas, N., Johansson, H. K. L., Panagiotou, E. M., Holmer, M. L., Elmelund, E., Zilliacus, J., Beronius, A., Damdimopoulou, P., van Duursen, M., & Svingen, T. (2024). AOP Report: An Upstream Network for Reduced Androgen Signaling Leading to Altered Gene Expression of Androgen Receptor–Responsive Genes in Target Tissues. Environmental Toxicology and Chemistry, 43(11), 2329–2337. https://doi.org/10.1002/etc.5972
Goldman AS, Shapiro B, & Neumann F. (1976). Role of testosterone and its metabolites in the differentiation of the mammary gland in rats. Endocrinology, 99(6), 1490–1495. https://doi.org/10.1210/endo-99-6-1490
Holmer ML, Zilliacus J, Draskau MK, Hlisníková H, Beronius A, Svingen T. Methodology for developing data-rich Key Event Relationships for Adverse Outcome Pathways exemplified by linking decreased androgen receptor activity with decreased anogenital distance. Reprod Toxicol. 2024 Sep;128:108662. doi: 10.1016/j.reprotox.2024.108662 . Epub 2024 Jul 8. PMID: 38986849.
Imperato-McGinley J, Binienda Z, Gedney J, & Vaughan ED Jr. (1986). Nipple differentiation in fetal male rats treated with an inhibitor of the enzyme 5 alpha-reductase: definition of a selective role for dihydrotestosterone. Endocrinology, 118(1), 132–137. https://doi.org/10.1210/endo-118-1-132
Knapczyk-Stwora, K., Nynca, A., Ciereszko, R. E., Paukszto, L., Jastrzebski, J. P., Czaja, E., Witek, P., Koziorowski, M., & Slomczynska, M. (2019). Flutamide-induced alterations in transcriptional profiling of neonatal porcine ovaries. Journal of Animal Science and Biotechnology, 10(1), 35. https://doi.org/10.1186/s40104-019-0340-y
Kratochwil, K., & Schwartz, P. (1976). Tissue interaction in androgen response of embryonic mammary rudiment of mouse: identification of target tissue for testosterone. Proceedings of the National Academy of Sciences, 73(11), 4041–4044. https://doi.org/10.1073/pnas.73.11.4041
Leung, J. K., & Sadar, M. D. (2017). Non-Genomic Actions of the Androgen Receptor in Prostate Cancer. Frontiers in Endocrinology, 8. https://doi.org/10.3389/fendo.2017.00002
Mendonca, B. B., Batista, R. L., Domenice, S., Costa, E. M. F., Arnhold, I. J. P., Russell, D. W., & Wilson, J. D. (2016). Steroid 5α-reductase 2 deficiency. The Journal of Steroid Biochemistry and Molecular Biology, 163, 206–211. https://doi.org/10.1016/j.jsbmb.2016.05.020
OECD (2008), Guidance Document on Mammalian Reproductive Toxicity Testing and Assessment, OECD Series on Testing and Assessment, No. 43, OECD Publishing, Paris, https://doi.org/10.1787/d2631d22-en.
OECD (2013), Guidance Document Supporting OECD Test Guideline 443 on the Extended One-Generational Reproductive Toxicity Test, OECD Series on Testing and Assessment, No. 151, OECD Publishing, Paris, ENV/JM/MONO(2013)10
OECD. (2023). Test No. 458: Stably Transfected Human Androgen Receptor Transcriptional Activation Assay for Detection of Androgenic Agonist and Antagonist Activity of Chemicals. OECD Publishing. https://doi.org/10.1787/9789264264366-en
OECD. (2025a). Test No. 421: Reproduction/Developmental Toxicity Screening Test. https://doi.org/10.1787/9789264264380-en
OECD. (2025b). Test No. 422: Combined Repeated Dose Toxicity Study with the Reproduction/Developmental Toxicity Screening Test. https://doi.org/10.1787/9789264264403-en
OECD. (2025c). Test No. 443: Extended One-Generation Reproductive Toxicity Study. https://doi.org/10.1787/9789264185371-en
Ogino, Y., Ansai, S., Watanabe, E. et al. Evolutionary differentiation of androgen receptor is responsible for sexual characteristic development in a teleost fish. Nat Commun 14, 1428 (2023). https://doi.org/10.1038/s41467-023-37026-6
Pedersen, E. B., Christiansen, S., & Svingen, T. (2022). AOP key event relationship report: Linking androgen receptor antagonism with nipple retention. In Current Research in Toxicology (Vol. 3). Elsevier B.V. https://doi.org/10.1016/j.crtox.2022.100085
Robitaille, J., & Langlois, V. S. (2020). Consequences of steroid-5α-reductase deficiency and inhibition in vertebrates. General and Comparative Endocrinology, 290, 113400. https://doi.org/10.1016/j.ygcen.2020.113400
Schwartz, C. L., Christiansen, S., Hass, U., Ramhøj, L., Axelstad, M., Löbl, N. M., & Svingen, T. (2021). On the Use and Interpretation of Areola/Nipple Retention as a Biomarker for Anti-androgenic Effects in Rat Toxicity Studies. In Frontiers in Toxicology (Vol. 3). Frontiers Media S.A. https://doi.org/10.3389/ftox.2021.730752
Svingen, T., Villeneuve, D. L., Knapen, D., Panagiotou, E. M., Draskau, M. K., Damdimopoulou, P., & O’Brien, J. M. (2021). A Pragmatic Approach to Adverse Outcome Pathway Development and Evaluation. Toxicological Sciences, 184(2), 183–190. https://doi.org/10.1093/toxsci/kfab113
Tut, T. G., Ghadessy, F. J., Trifiro, M. A., Pinsky, L., & Yong, E. L. (1997). Long Polyglutamine Tracts in the Androgen Receptor Are Associated with Reduced Trans -Activation, Impaired Sperm Production, and Male Infertility 1. The Journal of Clinical Endocrinology & Metabolism, 82(11), 3777–3782. https://doi.org/10.1210/jcem.82.11.4385
Williams, A. J., Grulke, C. M., Edwards, J., McEachran, A. D., Mansouri, K., Baker, N. C., Patlewicz, G., Shah, I., Wambaugh, J. F., Judson, R. S., & Richard, A. M. (2017). The CompTox Chemistry Dashboard: a community data resource for environmental chemistry. Journal of Cheminformatics, 9(1), 61. https://doi.org/10.1186/s13321-017-0247-6
Wolf, C., Lambright, C., Mann, P., Price, M., Cooper, R. L., Ostby, J., & Earl Gray, L. J. (1999). Administration of potentially antiandrogenic pesticides (procymidone, linuron, iprodione, chlozolinate, p,p-DDE, and ketoconazole) and toxic substances (dibutyl-and diethylhexyl phthalate, PCB 169, and ethane dimethane sulphonate) during sexual differentiation produces diverse profiles of reproductive malformations in the male rat. Toxicology and Industrial Health, 15, 94–118. www.stockton-press.co.uk
You, L., Casanova, M., Archibeque-Engle, S., Sar, M., Fan, L.-Q., & Heck, A. (1998). Impaired Male Sexual Development in Perinatal Sprague-Dawley and Long-Evans Hooded Rats Exposed in Utero and Lactationally to p,p’-DDE. In TOX1COLOGICAL SCIENCES (Vol. 45). https://academic.oup.com/toxsci/article/45/2/162/1653877