
This Key Event Relationship is licensed under the Creative Commons BY-SA license. This license allows reusers to distribute, remix, adapt, and build upon the material in any medium or format, so long as attribution is given to the creator. The license allows for commercial use. If you remix, adapt, or build upon the material, you must license the modified material under identical terms.
Relationship: 3348
Title
Decrease, AR activation leads to nipple retention, increased
Upstream event
Downstream event
Key Event Relationship Overview
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding | Point of Contact | Author Status | OECD Status |
|---|---|---|---|---|---|---|
| 5α-reductase inhibition leading to increased nipple retention (NR) in male (rodent) offspring | non-adjacent | High | Terje Svingen (send email) | Under development: Not open for comment. Do not cite | ||
| Androgen receptor (AR) antagonism leading to nipple retention (NR) in male (mammalian) offspring | non-adjacent | High | Terje Svingen (send email) | Under development: Not open for comment. Do not cite | Under Development | |
| Decreased testosterone synthesis leading to increased nipple retention (NR) in male (rodent) offspring | non-adjacent | High | Terje Svingen (send email) | Under development: Not open for comment. Do not cite |
Taxonomic Applicability
Sex Applicability
| Sex | Evidence |
|---|---|
| Male | High |
Life Stage Applicability
| Term | Evidence |
|---|---|
| Foetal | High |
Key Event Relationship Description
This KER links a decrease in androgen receptor (AR) activation during fetal development to increased nipple/areola retention (NR) in male rodent offspring. It should be noted that the upstream Key Event (KE) ‘decrease, androgen receptor activation’ (KE-1614 in AOP Wiki) specifically focuses on decreased activation of the AR in vivo, while most methods that can be used to measure AR activity are carried out in vitro. Indirect information about this KE may, for example, be provided from assays showing in vitro AR antagonism, decreased in vitro or in vivo testosterone production/levels, or decreased in vitro or in vivo dihydrotestosterone (DHT) production/levels.
The KER is not directly applicable to humans as both sexes have two nipples, and there is no known effect of androgens on their development (Schwartz et al., 2021). However, NR is a clear readout of reduced androgen action, impaired fetal masculinisation and disrupted sexual differentiation during development, which is relevant to humans, mammals (Schwartz et al., 2021), and vertebrates more broadly (Ogino et al., 2023). It is included as a mandatory endpoint in several rodent OECD Test Guidelines (OECD, 2025a, 2025b, 2025c) and, in OECD GD 151, is considered an adverse outcome applicable to the setting of Points of Departure for use in human health risk assessment (OECD, 2013). NR can also be used as an indicator of anti-androgenicity in mammals and vertebrates in the environment due to the conserved nature of the AR and its implication in sexual differentiation across species (Ogino et al., 2023).
Evidence Collection Strategy
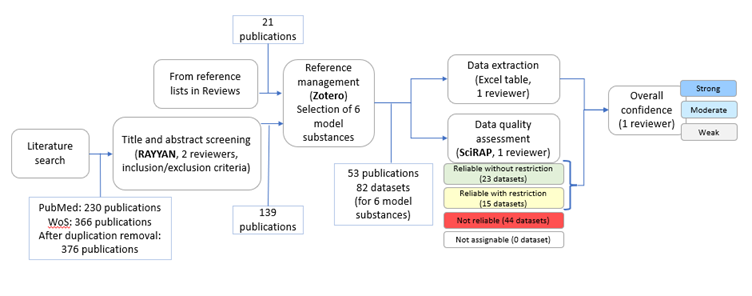
A systematic weight-of-evidence approach was applied to collect, evaluate, extract, and integrate evidence from in vivo studies (Figure 1, 71z732d7be_KER_3348_figure_1.png (752×296)).
{kind=link}
Search terms and search strings were developed for PubMed and Web of Science.
PubMed: ("androgen receptor*" OR "testosterone receptor*" OR "receptors, androgen"[MeSH Terms] OR "androgen*" OR "testosterone*" OR "dihydrotestosterone*" OR "androgens"[MeSH Terms] OR "androgen antagonists"[MeSH Terms]) AND ("nipple*" OR "areola*")
Web of Science: ("androgen receptor*" OR "testosterone receptor*" OR "androgen*" OR "testosterone*" OR "dihydrotestosterone*") AND ("nipple*" OR "areola*")
Literature searches in PubMed and Web of Science were performed on January 16, 2024. The obtained publications were imported into RAYYAN software (https://www.rayyan.ai) and duplicates were removed, resulting in 376 publications (Figure 1).
The following inclusion and exclusion criteria were used for screening the titles and abstracts in RAYYAN.
Inclusion criteria:
- Primary literature on
- exposure to androgenic or antiandrogenic compounds and nipple/areola retention as an outcome in in vivo studies in mammals
- measurement of androgen levels, AR activity or other androgen biomarkers, and nipple/areola retention in in vivo studies in mammals
- In vitro and in vivo mechanistic studies on nipple/areola retention
- Reviews on nipple/areola retention
Exclusion criteria:
- Not in English
- Abstracts and other non-full-text publications
139 publications were included based on the screening of the titles and abstracts by two reviewers. In addition, 21 publications from the reference lists of two previous reviews on NR were included (Pedersen et al., 2022; Schwartz et al., 2021).
Data extraction and study quality assessment for in vivo studies on nipple/areola retention
Information on six model substances was included to cover different mechanisms leading to reduced androgen receptor (AR) activation, including AR antagonism and decreased production of testosterone or dihydrotestosterone (DHT). The model substances were: DEHP, DBP, finasteride, flutamide, procymidone, or vinclozolin. Thus, full-text publications on in vivo studies of effects on NR in mammals after prenatal exposure to DEHP, DBP, finasteride, flutamide, procymidone, or vinclozolin were analysed by one reviewer per model substance. In short, data from the publications was extracted into an Excel template. Studies were divided into different datasets if different rat strains, exposure windows or time points for observation of NR were used. The reliability of the datasets was assessed using the SciRAP tool (http://www.scirap.org). In addition to the guidance for evaluation of each methodological quality criterion available in the SciRAP tool, specific considerations were developed and applied in this case and are listed in Table 1 in Appendix 1 (56ltlyy5fk_KER_3348_Appendix_1.pdf).
For each dataset, the result of the SciRAP evaluation was translated into reliability categories using the principles laid out in Table 1.
Table 1. Principles for translation of SciRAP evaluation results to reliability categories.
|
Reliability Category |
Principles |
|
1. Reliable without restriction |
SciRAP methodological quality Score > 80 and all key criteria* are “Fulfilled” and there are no deficiencies in the non-key criteria that might affect study reliability. |
|
2. Reliable with restrictions |
SciRAP methodological quality Score > 65 and one or several of the key criteria are “Partially Fulfilled”, or there are minor deficiencies in the non-key criteria that might affect study reliability. |
|
3. Not reliable |
SciRAP methodological quality Score < 65 or one or several of the key criteria are “Not Fulfilled”, or there are major deficiencies in the non-key criteria that affect reliability. |
|
4. Not assignable |
Two or more of the key criteria are “Not Determined” |
*Key criteria are criteria judged as specifically critical for the reliability of the data in a certain case and are determined a priori. The following five SciRAP criteria for methodological quality were considered as key criteria: A concurrent negative control group was included; The timing and duration of administration were appropriate for investigating the included endpoints; Measurements were collected at suitable time points in order to generate sensitive, valid, and reliable data; A sufficient number of animals per dose group were subjected to separate tests/data collection/measurements to generate reliable and valid results; and The statistical methods have been clearly described and do not seem inappropriate, unusual or unfamiliar.
The level of confidence in the overall data for each substance was categorised using the principles laid out in Table 2.
Table 2. Principles for categorisation of the level of confidence.
|
Level of confidence |
Principles for Categorisation** |
|
Strong |
OR
|
|
Moderate |
|
|
Weak |
OR
|
|
No effect |
|
**Conflicting results from datasets judged as not reliable did not impact the categorisation.
Identification of studies on the mechanistic understanding of substances causing increased nipple/areola retention
Relevant studies describing mechanisms for the substances flutamide, procymidone, vinclozolin, finasteride and DEHP were derived from the empirical evidence for the shared upstream KE decreased AR activation described recently for KER 2820 (Holmer et al., 2024). For DBP, which was not used as a model substance in KER 2820, studies describing mechanisms were identified from 1) the publications included on the in vivo studies of DBP in this KER and 2) publications on mechanisms of DEHP and DBP included in (Holmer et al., 2024).
Evidence Supporting this KER
Biological Plausibility
The biological plausibility for this KER is judged to be high based on the following:
- Sexual differentiation happens in fetal life. The testes are developed and start to produce testosterone that is converted in other tissues by the enzyme 5-alpha-reductase to the more potent androgen dihydrotestosterone (DHT). Both hormones bind and activate the nuclear receptor and transcription factor AR, which in turn drives the masculinization of the male fetus (Schwartz et al., 2021; Welsh et al., 2014).
- Fetal masculinization depends on the activation of androgen signalling during a critical time window, the masculinization programming window (MPW), which is observed around, embryonic day 14.5-17.5 in mice (Amato et al., 2022), gestational day (GD) 16-20 in rats and presumably gestation weeks (GWs) 8-14 in humans (Welsh et al., 2008).
- The fetal masculinization process involves a range of tissues and organs, including the nipple anlagen in rats and mice. In humans, both sexes have two nipples. In contrast, common laboratory mice and rats are sexually dimorphic, with females having 12 (rats) and 10 (mice) nipples and males generally having none (Mayer et al., 2008; Schwartz et al., 2021). In both male and female mouse embryos, stem cells differentiate into a mammary gland, with nipple anlagen being visible by embryonic day 11.5 (Mayer et al., 2008). In male embryos, the presence of androgen leads the nipple anlagen to regress a few days later (Kratochwil, 1977; Kratochwil & Schwartz, 1976) . The androgen responsiveness in the nipple anlagen is rather short, in mice starting late embryonic day 13, with loss of responsiveness on embryonic day 15 (Imperato-McGinley et al., 1986; Kratochwil, 1977) and thus roughly following the timing of the MPW.
- Nipple formation is inhibited in female mice and rat fetuses exposed to androgens during gestation (Goldman et al., 1976; Greene et al., 1941; Imperato-McGinley et al., 1986).
- Male Tfm-mutant mice, which are insensitive to androgens and believed to be so due to a nonfunctional androgen receptor, present with retained nipples (Kratochwil & Schwartz, 1976).
- Multiple mechanisms of action may potentially lead to nipple retention in male mouse and rat offspring. DHT is the main androgen responsible for nipple/areola regression through interaction with AR in the nipple anlagen (Imperato-McGinley et al., 1986). Inhibition of testosterone synthesis or inhibition of DHT to testosterone conversion, increased metabolism of androgens, or direct interference with AR activation may thus all lead to nipple/areola retention (Imperato-McGinley et al., 1986; Schwartz et al., 2021).
Empirical Evidence
The empirical support from studies in animals for this KER is judged as high overall.
It should be noted that the KE decreased AR activation (KE 1614 in AOP Wiki) specifically focuses on decreased activation of the AR in vivo, with no methods currently available to measure this. Examples of assays that provide indirect information about KE 1614 are described in upstream MIE/KEs.
The empirical evidence for this KER from animal studies in vivo is based on studies using six different substances that result in decreased AR activation by different mechanisms. Flutamide, procymidone and vinclozolin bind to the AR and inhibit the receptor activity and thereby act as AR antagonists, see MIE 26. Finasteride inhibits the 5-alpha-reductase enzyme that converts testosterone to DHT, see MIE 1617. DEHP and DBP exposure during prenatal development in rats results in reduced fetal testosterone levels, see KE-2298 and KE1690. (MIE 26, MIE 1617 and KE 1690 can be found in AOP-Wiki).
The evidence for the upstream KE is mainly based on data from in vitro assays (AR antagonism or 5-alpha-reductase inhibition in vitro), whereas the evidence for the downstream KE is based on in vivo studies, and there is generally no evidence for both KEs from the same study. However, decreased testosterone levels can be measured in vivo, and (Howdeshell et al., 2007; Martino-Andrade et al., 2009) measured the effect of developmental phthalate exposure on both testosterone levels and nipple/areola retention (see the section about “Dose concordance”).
The empirical evidence for the six substances is summarised in Table 3.
Table 3. Summary of empirical evidence for decreased androgen receptor activation, leading to increased nipple/areola retention. References for the studies supporting the empirical evidence are found in the section “Evidence for decreased AR activation (KE 1614) by flutamide, procymidone, and vinclozolin, finasteride, DEHP and DBP” and in Table 4 in Appendix 2 (5yw914oi7s_KER_3348_Appendix_2.pdf).
|
Stressor(s) |
Upstream effect (decreased AR activation) |
Downstream effect (Increased nipple/areola retention) |
|
Flutamide |
AR antagonism in in vitro assay, receptor binding and transactivation assays |
Increased NR in males after prenatal exposure in studies in rat |
|
Procymidone |
AR antagonism in in vitro assay receptor binding and transactivation assays |
Increased NR in males after prenatal exposure in studies in rat |
|
Vinclozolin |
AR antagonism in in vitro assay receptor binding and transactivation assays |
Increased NR in males after prenatal exposure in studies in rat |
|
Finasteride |
Inhibition of 5-alpha-reductase enzyme in in vitro assays |
Increased NR in males after prenatal exposure in studies in rat |
|
DEHP |
Reduced production of testosterone in fetal testis measured in ex vivo testis assays, reduced testosterone levels in testis, and reduced fetal plasma or serum testosterone levels |
Increased NR in males after prenatal exposure in studies in rat |
|
DBP |
Reduced production of testosterone in fetal testis measured in ex vivo testis assays and reduced testosterone levels in fetal testis |
Increased NR in males after prenatal exposure in studies in rat |
From Table 3, it can be deduced that fetal exposure to substances known to decrease androgen receptor activation through antagonism of the AR (vinclozolin, procymidone, flutamide), inhibition of testosterone synthesis (DEHP, DBP) or inhibition of the conversion of testosterone to DHT (finasteride), results in increased nipple/areola retention in rat male offspring.
Evidence for decreased AR activation (KE 1614) by flutamide, procymidone, vinclozolin, finasteride, DEHP and DBP.
Flutamide, a pharmaceutical, binds the AR and inhibits its activity, thereby acting as an AR antagonist. It has been used as an antiandrogen for the treatment of prostate cancer and is used as a reference chemical for antiandrogenic activity in the AR transactivation assays in the OECD test guideline No 458 (Goldspiel & Kohler, 1990; Labrie, 1993; OECD, 2023; Simard et al., 1986)
Procymidone and vinclozolin are fungicides that have been shown to be AR antagonists. Procymidone binds to the AR and inhibits the agonist binding, as shown in AR binding assays using rat prostate cytosol (Hosokawa et al., 1993) or AR transfected cells (Ostby et al., 1999). Procymidone also inhibits agonist activated transcription in AR reporter assays (Hass et al., 2012; Kojima et al., 2004; Orton et al., 2011; Ostby et al., 1999; Scholze et al., 2020). Vinclozolin binds to the AR and inhibits the agonist binding, as shown in AR binding assays using rat epididymis cytosol (Kelce & Wilson, 1997) or AR transfected cells (Wong et al., 1995). Vinclozolin also inhibits agonist activated transcription in AR reporter assays (Euling, 2002; Kojima et al., 2004; Molina-Molina et al., 2006; Orton et al., 2011; Scholze et al., 2020; Shimamura et al., 2002; Wong et al., 1995).
Finasteride is a pharmaceutical that inhibits the 5-alpha-reductase enzyme that converts testosterone to DHT. Finasteride is used to treat benign prostatic hypertrophy (Andersson & Russell, 1990; Stoner, 1990; Wood & Rittmaster, 1994).
Prenatal exposure to DEHP in rats has been shown to reduce the production of testosterone in fetal testis measured in ex vivo testis assays, and to reduce testosterone levels in testis and in fetal plasma and serum (Borch et al., 2006; Borch J et al., 2004; Culty et al., 2008; Hannas et al., 2011, 2012; Howdeshell et al., 2007; Klinefelter et al., 2012; Parks, 2000; VO et al., 2009; Wilson et al., 2004, 2007). Conversely, prenatal DEHP exposure did not result in any effects on testosterone levels in the testis at PND1 in one study by Andrade et al. (2006) (Andrade et al., 2006). Similar to DEHP, prenatal exposure to DBP has been shown to reduce the production of testosterone in fetal rat testis measured in ex vivo testis studies (Howdeshell et al., 2007; Wilson et al., 2004) and reduce testosterone levels in the fetal rat testis (Martino-Andrade et al., 2009). The precise underlying mechanism for these effects of DEHP and DPB is presently unknown.
Evidence for increased nipple/areola retention in males (AO-1786) by prenatal exposure to flutamide, procymidone, vinclozolin, finasteride, DEHP and DBP.
All datasets that were used for the weight of evidence assessment were judged as reliable without or with restriction. The majority of datasets assessed showed an increased nipple/areola retention in male offspring after gestational exposure. The conclusion was that the level of confidence was strong for all six substances. The studies are summarised in Table 4 in Appendix 2, 5yw914oi7s_KER_3348_Appendix_2.pdf
Dose concordance
Dose concordance is challenging to assess for this KER since in vivo AR activity is currently not possible to measure but can only be inferred indirectly by measures of upstream events. In some studies, fetal (testicular) testosterone levels during, or close to, the fetal masculinization programming window are measured in a subset of animals exposed similarly to those investigated for NR postnatally. Such information may inform on dose concordance if more doses are included.
In a rat in utero exposure study (GD13-21) with DPB and DEHP, testosterone levels in the fetal testes were investigated at GD21, and NR was investigated at PND13 (Martino-Andrade et al., 2009). For DBP, both reduced testosterone levels in fetal testes and NR were observed at 500 mg/kg/d, whereas no effect on NR and only a slight non-significant reduction of testosterone was observed at the lower dose (100 mg/kg/d). For DEHP, a slight but non-significant decrease in testosterone levels in fetal rat testis was observed after exposure to 150 mg/kg/d DEHP, with no effects on nipple/areola retention.
Such data could suggest dose concordance for this part of the KER, although the evidence for this is not strong.
Temporal concordance
Temporal concordance can only be considered from a theoretical perspective since the downstream event, increased NR, is a result of disruption to normal regression of nipple anlagen in male rodents induced during a short window of gestational development (in mice of approximately 2 days) but usually measured at PND12-14 in rats. Earlier than this, the areolae are not yet visible through the skin and later than this, the animals grow fur and need to be shaved for proper examination. This is supported by several of the studies in the empirical evidence, where the test substance was administered during a short period during gestation and nipple retention was observed postnatally.
Based on current knowledge, it is understood that the upstream event – decreased AR activation in vivo – takes place minutes to hours after exposure to an anti-androgenic substance. If a substance decreases AR activation through inhibition of the AR, the upstream event is expected to happen immediately after exposure. If a substance decreases androgen receptor activation through inhibition of testosterone synthesis, the upstream event is expected to happen minutes to hours after the exposure.
Uncertainties and Inconsistencies
For DEHP and DBP, there were some inconsistencies in the empirical evidence, but they could be explained by differences in study designs and uncertainties in measurements (see Appendix 1). Some uncertainty is imposed by the poorly supported dose-concordance. However, the dose-concordance is well supported by the current understanding of biological processes.
Known modulating factors
A well established modulating factor for androgen action is genetic variations in the AR, which decrease the function of the receptor. For example, longer CAG repeat lengths have been associated with decreased AR activation (Chamberlain et al., 1994; Tut et al., 1997). Rat strain is another important modulating factor, with studies showing that the Long-Evans Hooded rat is less sensitive to nipple/areola retention than the Sprague-Dawley rat (Wolf et al., 1999; You et al., 1998)
Quantitative Understanding of the Linkage
The quantitative understanding of the linkage is low. This is a consequence of it not being possible to measure the upstream and the downstream events in the same study.
Response-response Relationship
The difficulties in extrapolating potency from in vitro to in vivo studies were exemplified by a comparison of the effects of pyrifluquinazon and bisphenol C in vitro and in utero. In vitro, bisphenol C antagonized the androgen receptor with a much higher potency than pyrifluquinazon, but in vivo the potencies were reversed with pyrifluquinazon exposure leading to NR at lower exposure levels than bisphenol C (Gray et al., 2019).
Time-scale
AR activation operates on a time-scale of minutes. The AR is a ligand-activated nuclear receptor and transcription factor. Upon ligand binding a conformational change and subsequent dimerization of the AR takes place within 3-6 minutes (Schaufele et al., 2005). Nuclear translocation (Nightingale et al., 2003) and promoter interactions occur within 15 minutes of ligand binding, and RNA polymerase II and coactivator recruitment are then proposed to occur transiently with cycles of approximately 90 minutes (Kang et al., 2002).
For the downstream event, the time-scale for observing a measurable effect on nipple/areola retention is closer to days and weeks, depending on species. For instance, in mice the nipple anlage are responsive to androgen action at embryonic day 13-15, while a sexual dimorphism of the nipples/areolas can first be observed after birth (Imperato-McGinley et al., 1986) .
Known Feedforward/Feedback loops influencing this KER
Not relevant for this KER.
Domain of Applicability
The KER is considered directly applicable to rats and mice, in which males normally have no nipples due to high levels of androgens during development, leading to regression of nipple anlagen. The empirical evidence supports the relevance to rats, whereas the relevance in mice is assumed based on knowledge about developmental biology in this species. Applicability may extend to most rodents.
While NR is not directly translatable to humans, it serves as a clear indicator of diminished androgen activity causing disrupted fetal masculinisation and sexual differentiation during development - an effect considered relevant to mammals, humans (Schwartz et al., 2021), and vertebrates more broadly (Ogino et al., 2023). NR is included as a mandatory endpoint in several rodent OECD Test Guidelines (OECD 2025a; OECD 2025b, OECD 2025c) and in OECD GD 151 considered an adverse outcome applicable to the setting of Points of Departure for use in human health risk assessment (OECD, 2013). NR can also be used as an indicator of anti-androgenicity in mammals and vertebrates in the environment due to the conserved nature of the AR and its implication in sexual differentiation across species (Ogino et al., 2023).
Life stage
Programming of nipple/areola regression in males occurs during a short window of sensitivity to androgens in the nipple anlagen during fetal life. This takes place in rats around embryonic days 13-15 (Imperato-McGinley et al., 1986), which is, therefore, the relevant window of exposure. The relevant timing for the investigation of NR is PND12-14 in male rat offspring when the nipples are visible in the female littermates. At this time in development, the nipples/areolas are visible through the skin without excessive fur that may interfere with the investigation (Schwartz et al., 2021). It should be mentioned that though the occurrence of nipples/areolas in male offspring is believed to be relatively stable throughout life, it may be responsive to postnatal changes. Permanent nipple/areola retention is observed in some but not all in utero exposure studies with antiandrogens inducing nipple/areola retention at PND 12-14. Most of the differences between studies seem explainable by the window of exposure, dose levels and methods for investigation used, but the responsiveness of nipple/areola retention to postnatal changes remains to be fully explored (Schwartz et al., 2021).
Sex
Data presented in this KER support that disruption of androgen action during fetal life can lead to increased nipple/areola retention in male rat offspring. Since female mice and rat offspring, in general, have 10 (mice) or 12 (rats) nipples at the relevant time of investigation, increased nipple/areola retention at that time point is not a relevant endpoint for females.
References
Amato, C. M., Yao, H. H.-C., & Zhao, F. (2022). One Tool for Many Jobs: Divergent and Conserved Actions of Androgen Signaling in Male Internal Reproductive Tract and External Genitalia. Frontiers in Endocrinology, 13. https://doi.org/10.3389/fendo.2022.910964
Andersson, S., & Russell, D. W. (1990). Structural and biochemical properties of cloned and expressed human and rat steroid 5 alpha-reductases. Proceedings of the National Academy of Sciences, 87(10), 3640–3644. https://doi.org/10.1073/pnas.87.10.3640
Andrade, A. M., Grande, S. W., Talsness, C. E., Grote, K., & Chahoud, I. (2006). Developmental exposure to high and low doses of di-(2-ethylhexyl) phthalate (DEHP); Effects on male rat offspring. NAUNYN-SCHMIEDEBERGS ARCHIVES OF PHARMACOLOGY, 372, 100–101.
Barlow, N. J., McIntyre, B. S., & Foster, P. M. D. (2004). Male reproductive tract lesions at 6, 12, and 18 months of age following in utero exposure to di(n-butyl) phthalate. TOXICOLOGIC PATHOLOGY, 32(1), 79–90. https://doi.org/10.1080/01926230490265894
Borch, J., Axelstad, M., Vinggaard, A. M., & Dalgaard, M. (2006). Diisobutyl phthalate has comparable anti-androgenic effects to di-n-butyl phthalate in fetal rat testis. Toxicology Letters, 163(3), 183–190. https://doi.org/10.1016/j.toxlet.2005.10.020
Borch J, Ladefoged O, Hass U, & Vinggaard AM. (2004). Steroidogenesis in fetal male rats is reduced by DEHP and DINP, but endocrine effects of DEHP are not modulated by DEHA in fetal, prepubertal and adult male rats. Reproductive Toxicology (Elmsford, N.Y.), 18(1), 53–61. https://doi.org/10.1016/j.reprotox.2003.10.011
Chamberlain, N. L., Driver, E. D., & Miesfeld, R. L. (1994). The length and location of CAG trinucleotide repeats in the androgen receptor N-terminal domain affect transactivation function. Nucleic Acids Research, 22(15), 3181–3186. https://doi.org/10.1093/nar/22.15.3181
Christiansen S, Scholze M, Dalgaard M, Vinggaard AM, Axelstad M, Kortenkamp A, & Hass U. (2009). Synergistic disruption of external male sex organ development by a mixture of four antiandrogens. Environmental Health Perspectives, 117(12), 1839–1846. https://doi.org/10.1289/ehp.0900689
Christiansen S, Boberg J, Axelstad M, Dalgaard M, Vinggaard AM, Metzdorff SB, & Hass U. (2010). Low-dose perinatal exposure to di(2-ethylhexyl) phthalate induces anti-androgenic effects in male rats. Reproductive Toxicology (Elmsford, N.Y.), 30(2), 313–321. https://doi.org/10.1016/j.reprotox.2010.04.005
Clewell, R. A., Thomas, A., Willson, G., Creasy, D. M., & Andersen, M. E. (2013). A dose response study to assess effects after dietary administration of diisononyl phthalate (DINP) in gestation and lactation on male rat sexual development. REPRODUCTIVE TOXICOLOGY, 35, 70–80. https://doi.org/10.1016/j.reprotox.2012.07.008
Culty, M., Thuillier, R., Li, W., Wang, Y., Martinez-Arguelles, D. B., Benjamin, C. G., Triantafilou, K. M., Zirkin, B. R., & Papadopoulos, V. (2008). In Utero Exposure to Di-(2-ethylhexyl) Phthalate Exerts Both Short-Term and Long-Lasting Suppressive Effects on Testosterone Production in the Rat1. Biology of Reproduction, 78(6), 1018–1028. https://doi.org/10.1095/biolreprod.107.065649
Euling, S. Y. (2002). Response-Surface Modeling of the Effect of 5alpha-Dihydrotestosterone and Androgen Receptor Levels on the Response to the Androgen Antagonist Vinclozolin. Toxicological Sciences, 69(2), 332–343. https://doi.org/10.1093/toxsci/69.2.332
Fussell KC, Schneider S, Buesen R, Groeters S, Strauss V, Melching-Kollmuss S, & van Ravenzwaay B. (2015). Investigations of putative reproductive toxicity of low-dose exposures to flutamide in Wistar rats. Archives of Toxicology, 89(12), 2385–2402. https://doi.org/10.1007/s00204-015-1622-6
Goldman AS, Shapiro B, & Neumann F. (1976). Role of testosterone and its metabolites in the differentiation of the mammary gland in rats. Endocrinology, 99(6), 1490–1495. https://doi.org/10.1210/endo-99-6-1490
Goldspiel, B. R., & Kohler, D. R. (1990). Flutamide: An Antiandrogen for Advanced Prostate Cancer. DICP, 24(6), 616–623. https://doi.org/10.1177/106002809002400612
Gray LE Jr, Ostby J, Furr J, Price M, Veeramachaneni DN, & Parks L. (2000). Perinatal exposure to the phthalates DEHP, BBP, and DINP, but not DEP, DMP, or DOTP, alters sexual differentiation of the male rat. Toxicological Sciences : An Official Journal of the Society of Toxicology, 58(2), 350–365. https://doi.org/10.1093/toxsci/58.2.350
Gray LE Jr, Barlow NJ, Howdeshell KL, Ostby JS, Furr JR, & Gray CL. (2009). Transgenerational effects of Di (2-ethylhexyl) phthalate in the male CRL:CD(SD) rat: added value of assessing multiple offspring per litter. Toxicological Sciences : An Official Journal of the Society of Toxicology, 110(2), 411–425. https://doi.org/10.1093/toxsci/kfp109
Gray, L. E., Furr, J. R., Conley, J. M., Lambright, C. S., Evans, N., Cardon, M. C., Wilson, V. S., Foster, P. M., & Hartig, P. C. (2019). A Conflicted Tale of Two Novel AR Antagonists in Vitro and in Vivo: Pyrifluquinazon Versus Bisphenol C. Toxicological Sciences, 168(2), 632–643. https://doi.org/10.1093/toxsci/kfz010
Greene, R. R., Burrill, M. W., & Ivy, A. C. (1941). Experimental intersexuality: The effects of combined estrogens and androgens on the embryonic sexual development of the rat. Journal of Experimental Zoology, 87(2), 211–232. https://doi.org/10.1002/jez.1400870203
Hannas, B. R., Furr, J., Lambright, C. S., Wilson, V. S., Foster, P. M. D., & Gray, L. E. (2011). Dipentyl Phthalate Dosing during Sexual Differentiation Disrupts Fetal Testis Function and Postnatal Development of the Male Sprague-Dawley Rat with Greater Relative Potency than Other Phthalates. Toxicological Sciences, 120(1), 184–193. https://doi.org/10.1093/toxsci/kfq386
Hannas, B. R., Lambright, C. S., Furr, J., Evans, N., Foster, P. M. D., Gray, E. L., & Wilson, V. S. (2012). Genomic Biomarkers of Phthalate-Induced Male Reproductive Developmental Toxicity: A Targeted RT-PCR Array Approach for Defining Relative Potency. Toxicological Sciences, 125(2), 544–557. https://doi.org/10.1093/toxsci/kfr315
Hass U, Scholze M, Christiansen S, Dalgaard M, Vinggaard AM, Axelstad M, Metzdorff SB, & Kortenkamp A. (2007). Combined exposure to anti-androgens exacerbates disruption of sexual differentiation in the rat. Environmental Health Perspectives, 115, 122–128. https://doi.org/10.1289/ehp.9360
Hass U, Boberg J, Christiansen S, Jacobsen PR, Vinggaard AM, Taxvig C, Poulsen ME, Herrmann SS, Jensen BH, Petersen A, Clemmensen LH, & Axelstad M. (2012). Adverse effects on sexual development in rat offspring after low dose exposure to a mixture of endocrine disrupting pesticides. Reproductive Toxicology (Elmsford, N.Y.), 34(2), 261–274. https://doi.org/10.1016/j.reprotox.2012.05.090
Hellwig J, van Ravenzwaay B, Mayer M, & Gembardt C. (2000). Pre- and postnatal oral toxicity of vinclozolin in Wistar and Long-Evans rats. Regulatory Toxicology and Pharmacology : RTP, 32(1), 42–50. https://doi.org/10.1006/rtph.2000.1400
Holmer, M. L., Zilliacus, J., Draskau, M. K., Hlisníková, H., Beronius, A., & Svingen, T. (2024). Methodology for developing data-rich Key Event Relationships for Adverse Outcome Pathways exemplified by linking decreased androgen receptor activity with decreased anogenital distance. Reproductive Toxicology, 128, 108662. https://doi.org/10.1016/j.reprotox.2024.108662
Hosokawa S, Murakami M, Ineyama M, Yamada T, Yoshitake A, Yamada H, Miyamoto J. The affinity of procymidone to androgen receptor in rats and mice. J Toxicol Sci. 1993 May;18(2):83-93. doi: 10.2131/jts.18.83. PMID: 8331696.
Howdeshell, K. L., Furr, J., Lambright, C. R., Rider, C. V., Wilson, V. S., & Gray, L. E. (2007). Cumulative Effects of Dibutyl Phthalate and Diethylhexyl Phthalate on Male Rat Reproductive Tract Development: Altered Fetal Steroid Hormones and Genes. Toxicological Sciences, 99(1), 190–202. https://doi.org/10.1093/toxsci/kfm069
Imperato-McGinley J, Binienda Z, Gedney J, & Vaughan ED Jr. (1986). Nipple differentiation in fetal male rats treated with an inhibitor of the enzyme 5 alpha-reductase: definition of a selective role for dihydrotestosterone. Endocrinology, 118(1), 132–137. https://doi.org/10.1210/endo-118-1-132
Jarfelt K, Dalgaard M, Hass U, Borch J, Jacobsen H, & Ladefoged O. (2005). Antiandrogenic effects in male rats perinatally exposed to a mixture of di(2-ethylhexyl) phthalate and di(2-ethylhexyl) adipate. Reproductive Toxicology (Elmsford, N.Y.), 19(4), 505–515. https://doi.org/10.1016/j.reprotox.2004.11.005
Kang, H.-Y., Huang, K.-E., Chang, S. Y., Ma, W.-L., Lin, W.-J., & Chang, C. (2002). Differential Modulation of Androgen Receptor-mediated Transactivation by Smad3 and Tumor Suppressor Smad4. Journal of Biological Chemistry, 277(46), 43749–43756. https://doi.org/10.1074/jbc.M205603200
Kelce, W. R., & Wilson, E. M. (1997). Environmental antiandrogens: Developmental effects, molecular mechanisms, and clinical implications. JOURNAL OF MOLECULAR MEDICINE-JMM, 75(3), 198–207. https://doi.org/10.1007/s001090050104
Klinefelter, G. R., Laskey, J. W., Winnik, W. M., Suarez, J. D., Roberts, N. L., Strader, L. F., Riffle, B. W., & Veeramachaneni, D. N. R. (2012). Novel molecular targets associated with testicular dysgenesis induced by gestational exposure to diethylhexyl phthalate in the rat: a role for estradiol. REPRODUCTION, 144(6), 747–761. https://doi.org/10.1530/REP-12-0266
Kojima, H., Katsura, E., Takeuchi, S., Niiyama, K., & Kobayashi, K. (2004). Screening for estrogen and androgen receptor activities in 200 pesticides by in vitro reporter gene assays using Chinese hamster ovary cells. Environmental Health Perspectives, 112(5), 524–531. https://doi.org/10.1289/ehp.6649
Kratochwil, K. (1977). Development and loss of androgen responsiveness in the embryonic rudiment of the mouse mammary gland. Developmental Biology, 61(2), 358–365. https://doi.org/10.1016/0012-1606(77)90305-0
Kratochwil, K., & Schwartz, P. (1976). Tissue interaction in androgen response of embryonic mammary rudiment of mouse: identification of target tissue for testosterone. Proceedings of the National Academy of Sciences, 73(11), 4041–4044. https://doi.org/10.1073/pnas.73.11.4041
Labrie, F. (1993). Mechanism of action and pure antiandrogenic properties of flutamide. Cancer, 72(12 Suppl), 3816–3827. https://doi.org/10.1002/1097-0142(19931215)72:12+<3816::aid-cncr2820721711>3.0.co;2-3
Martino-Andrade AJ, Morais RN, Botelho GG, Muller G, Grande SW, Carpentieri GB, Leão GM, & Dalsenter PR. (2009). Coadministration of active phthalates results in disruption of foetal testicular function in rats. International Journal of Andrology, 32(6), 704–712. https://doi.org/10.1111/j.1365-2605.2008.00939.x
Matsuura I, Saitoh T, Tani E, Wako Y, Iwata H, Toyota N, Ishizuka Y, Namiki M, Hoshino N, Tsuchitani M, Ikeda Y. Evaluation of a two-generation reproduction toxicity study adding endpoints to detect endocrine disrupting activity using lindane. J Toxicol Sci. (2005) Dec;30 Spec No.:135-161. doi: 10.2131/jts.30.s135. PMID: 16641539.
Mayer, J. A., Foley, J., De La Cruz, D., Chuong, C. M., & Widelitz, R. (2008). Conversion of the nipple to hair-bearing epithelia by lowering bone morphogenetic protein pathway activity at the dermal-epidermal interface. The American Journal of Pathology, 173(5), 1339–1348. https://doi.org/10.2353/AJPATH.2008.070920
McIntyre BS, Barlow NJ, & Foster PM. (2001). Androgen-mediated development in male rat offspring exposed to flutamide in utero: permanence and correlation of early postnatal changes in anogenital distance and nipple retention with malformations in androgen-dependent tissues. Toxicological Sciences : An Official Journal of the Society of Toxicology, 62(2), 236–249. https://doi.org/10.1093/toxsci/62.2.236
Molina-Molina JM, Hillenweck A, Jouanin I, Zalko D, Cravedi JP, Fernández MF, Pillon A, Nicolas JC, Olea N, Balaguer P. Steroid receptor profiling of vinclozolin and its primary metabolites. Toxicol Appl Pharmacol. 2006 Oct 1;216(1):44-54. doi: 10.1016/j.taap.2006.04.005. Epub 2006 Jun 5. PMID: 16750840.
Moore RW, Rudy TA, Lin TM, Ko K, & Peterson RE. (2001). Abnormalities of sexual development in male rats with in utero and lactational exposure to the antiandrogenic plasticizer Di(2-ethylhexyl) phthalate. Environmental Health Perspectives, 109(3), 229–237. https://doi.org/10.1289/ehp.01109229
Mylchreest E, Wallace DG, Cattley RC, & Foster PM. (2000). Dose-dependent alterations in androgen-regulated male reproductive development in rats exposed to Di(n-butyl) phthalate during late gestation. Toxicological Sciences : An Official Journal of the Society of Toxicology, 55(1), 143–151. https://doi.org/10.1093/toxsci/55.1.143
Nightingale, J., Chaudhary, K. S., Abel, P. D., Stubbs, A. P., Romanska, H. M., Mitchell, S. E., Stamp, G. W. H., & Lalani, E.-N. (2003). Ligand Activation of the Androgen Receptor Downregulates E-Cadherin-Mediated Cell Adhesion and Promotes Apoptosis of Prostatic Cancer Cells. Neoplasia, 5(4), 347–361. https://doi.org/10.1016/S1476-5586(03)80028-3
OECD (2013), Guidance Document Supporting OECD Test Guideline 443 on the Extended One-Generational Reproductive Toxicity Test, OECD Series on Testing and Assessment, No. 151, OECD Publishing, Paris, ENV/JM/MONO(2013)10
OECD (2023). Test No. 458: Stably Transfected Human Androgen Receptor Transcriptional Activation Assay for Detection of Androgenic Agonist and Antagonist Activity of Chemicals. OECD Guidelines for the Testing of Chemicals, Section 4, OECD Publishing, Paris, https://doi.org/10.1787/9789264264366-en
OECD (2025a), Test No. 443: Extended One-Generation Reproductive Toxicity Study, OECD Guidelines for the Testing of Chemicals, Section 4, OECD Publishing, Paris, https://doi.org/10.1787/9789264185371-en.
OECD (2025b), Test No. 421: Reproduction/Developmental Toxicity Screening Test, OECD Guidelines for the Testing of Chemicals, Section 4, OECD Publishing, Paris, https://doi.org/10.1787/9789264264380-en.
OECD (2025c), Test No. 422: Combined Repeated Dose Toxicity Study with the Reproduction/Developmental Toxicity Screening Test, OECD Guidelines for the Testing of Chemicals, Section 4, OECD Publishing, Paris, https://doi.org/10.1787/9789264264403-en.
Ogino, Y., Ansai, S., Watanabe, E. et al. Evolutionary differentiation of androgen receptor is responsible for sexual characteristic development in a teleost fish. Nat Commun 14, 1428 (2023). https://doi.org/10.1038/s41467-023-37026-6
Orton, F., Rosivatz, E., Scholze, M., & Kortenkamp, A. (2011). Widely Used Pesticides with Previously Unknown Endocrine Activity Revealed as in Vitro Antiandrogens. ENVIRONMENTAL HEALTH PERSPECTIVES, 119(6), 794–800. https://doi.org/10.1289/ehp.1002895
Ostby J, Monosson E, Kelce WR, & Gray LE Jr. (1999). Environmental antiandrogens: low doses of the fungicide vinclozolin alter sexual differentiation of the male rat. Toxicology and Industrial Health, 15(1), 48–64. https://doi.org/10.1177/074823379901500106
Parks, L. G. (2000). The Plasticizer Diethylhexyl Phthalate Induces Malformations by Decreasing Fetal Testosterone Synthesis during Sexual Differentiation in the Male Rat. Toxicological Sciences, 58(2), 339–349. https://doi.org/10.1093/toxsci/58.2.339
Pedersen, E. B., Christiansen, S., & Svingen, T. (2022). AOP key event relationship report: Linking androgen receptor antagonism with nipple retention. In Current Research in Toxicology (Vol. 3). Elsevier B.V. https://doi.org/10.1016/j.crtox.2022.100085
Saillenfait AM, Sabaté JP, & Gallissot F. (2008). Diisobutyl phthalate impairs the androgen-dependent reproductive development of the male rat. Reproductive Toxicology (Elmsford, N.Y.), 26(2), 107–115. https://doi.org/10.1016/j.reprotox.2008.07.006
Saillenfait AM, Sabaté JP, & Gallissot F. (2009). Effects of in utero exposure to di-n-hexyl phthalate on the reproductive development of the male rat. Reproductive Toxicology (Elmsford, N.Y.), 28(4), 468–476. https://doi.org/10.1016/j.reprotox.2009.06.013
Schaufele, F., Carbonell, X., Guerbadot, M., Borngraeber, S., Chapman, M. S., Ma, A. A. K., Miner, J. N., & Diamond, M. I. (2005). The structural basis of androgen receptor activation: Intramolecular and intermolecular amino–carboxy interactions. Proceedings of the National Academy of Sciences, 102(28), 9802–9807. https://doi.org/10.1073/pnas.0408819102
Schneider S, Kaufmann W, Strauss V, & van Ravenzwaay B. (2011). Vinclozolin: a feasibility and sensitivity study of the ILSI-HESI F1-extended one-generation rat reproduction protocol. Regulatory Toxicology and Pharmacology : RTP, 59(1), 91–100. https://doi.org/10.1016/j.yrtph.2010.09.010
Scholze, M., Taxvig, C., Kortenkamp, A., Boberg, J., Christiansen, S., Svingen, T., Lauschke, K., Frandsen, H., Ermler, S., Hermann, S. S., Pedersen, M., Lykkeberg, A. K., Axelstad, M., & Vinggaard, A. M. (2020). Quantitative in Vitro to in Vivo Extrapolation (QIVIVE) for Predicting Reduced Anogenital Distance Produced by Anti-Androgenic Pesticides in a Rodent Model for Male Reproductive Disorders. Environmental Health Perspectives, 128(11), 117005. https://doi.org/10.1289/EHP6774
Schwartz, C. L., Christiansen, S., Hass, U., Ramhøj, L., Axelstad, M., Löbl, N. M., & Svingen, T. (2021). On the Use and Interpretation of Areola/Nipple Retention as a Biomarker for Anti-androgenic Effects in Rat Toxicity Studies. In Frontiers in Toxicology (Vol. 3). Frontiers Media S.A. https://doi.org/10.3389/ftox.2021.730752
Shimamura M, Kodaira K, Kenichi H, Ishimoto Y, Tamura H, Iguchi T. Comparison of antiandrogenic activities of vinclozolin and D,L-camphorquinone in androgen receptor gene transcription assay in vitro and mouse in utero exposure assay in vivo. Toxicology. 2002 May 24;174(2):97-107. doi: 10.1016/s0300-483x(02)00044-6. PMID: 11985887.
Simard, J., Luthy, I., Guay, J., Bélanger, A., & Labrie, F. (1986). Characteristics of interaction of the antiandrogen flutamide with the androgen receptor in various target tissues. Molecular and Cellular Endocrinology, 44(3), 261–270. https://doi.org/10.1016/0303-7207(86)90132-2
Stoner, E. (1990). The clinical development of a 5α-reductase inhibitor, finasteride. The Journal of Steroid Biochemistry and Molecular Biology, 37(3), 375–378. https://doi.org/10.1016/0960-0760(90)90487-6
Test No. 458: Stably Transfected Human Androgen Receptor Transcriptional Activation Assay for Detection of Androgenic Agonist and Antagonist Activity of Chemicals. (2023). OECD. https://doi.org/10.1787/9789264264366-en
Tut, T. G., Ghadessy, F. J., Trifiro, M. A., Pinsky, L., & Yong, E. L. (1997). Long Polyglutamine Tracts in the Androgen Receptor Are Associated with Reduced Trans -Activation, Impaired Sperm Production, and Male Infertility 1. The Journal of Clinical Endocrinology & Metabolism, 82(11), 3777–3782. https://doi.org/10.1210/jcem.82.11.4385
Vo TT, Jung EM, Dang VH, Jung K, Baek J, Choi KC, Jeung EB. Differential effects of flutamide and di-(2-ethylhexyl) phthalate on male reproductive organs in a rat model. J Reprod Dev. 2009 Aug;55(4):400-11. doi: 10.1262/jrd.20220. Epub 2009 Apr 13. PMID: 19367084.
Welsh, M., Saunders, P. T. K., Fisken, M., Scott, H. M., Hutchison, G. R., Smith, L. B., & Sharpe, R. M. (2008). Identification in rats of a programming window for reproductive tract masculinization, disruption of which leads to hypospadias and cryptorchidism. JOURNAL OF CLINICAL INVESTIGATION, 118(4), 1479–1490. https://doi.org/10.1172/JCI34241
Welsh, M., Suzuki, H., & Yamada, G. (2014). The Masculinization Programming Window. In O. Hiort & S. F. Ahmed (Eds.), UNDERSTANDING DIFFERENCES AND DISORDERS OF SEX DEVELOPMENT (DSD) (Vol. 27, pp. 17–27). https://doi.org/10.1159/000363609
Wilson, V. S., Lambright, C., Furr, J., Ostby, J., Wood, C., Held, G., & Gray, L. E. (2004). Phthalate ester-induced gubernacular lesions are associated with reduced insl3 gene expression in the fetal rat testis. Toxicology Letters, 146(3), 207–215. https://doi.org/10.1016/j.toxlet.2003.09.012
Wilson, V. S., Howdeshell, K. L., Lambright, C. S., Furr, J., & Earl Gray, L. (2007). Differential expression of the phthalate syndrome in male SpragueDawley and Wistar rats after in utero DEHP exposure. Toxicology Letters, 170(3), 177–184. https://doi.org/10.1016/j.toxlet.2007.03.004
Wolf C Jr, Lambright C, Mann P, Price M, Cooper RL, Ostby J, Gray LE Jr. Administration of potentially antiandrogenic pesticides (procymidone, linuron, iprodione, chlozolinate, p,p'-DDE, and ketoconazole) and toxic substances (dibutyl- and diethylhexyl phthalate, PCB 169, and ethane dimethane sulphonate) during sexual differentiation produces diverse profiles of reproductive malformations in the male rat. Toxicol Ind Health. 1999 Jan-Mar;15(1-2):94-118. doi: 10.1177/074823379901500109. PMID: 10188194.
Wolf CJ, LeBlanc GA, Ostby JS, & Gray LE Jr. (2000). Characterization of the period of sensitivity of fetal male sexual development to vinclozolin. Toxicological Sciences : An Official Journal of the Society of Toxicology, 55(1), 152–161. https://doi.org/10.1093/toxsci/55.1.152
Wolf CJ, LeBlanc GA, & Gray LE Jr. (2004). Interactive effects of vinclozolin and testosterone propionate on pregnancy and sexual differentiation of the male and female SD rat. Toxicological Sciences : An Official Journal of the Society of Toxicology, 78(1), 135–143. https://pubmed.ncbi.nlm.nih.gov/14736997/
Wong, C. I., Kelce, W. R., Sar, M., & Wilson, E. M. (1995). Androgen Receptor Antagonist versus Agonist Activities on the Fungicide Vinclozolin Relative to Hydroxyflutamide. Nucleic Acids, Protein Synthesis, and Molecular Genetics, 270(34), 19998–20003.
Wood, A. J. J., & Rittmaster, R. S. (1994). Finasteride. New England Journal of Medicine, 330(2), 120–125. https://doi.org/10.1056/NEJM199401133300208
You, L., Casanova, M., Archibeque-Engle, S., Sar, M., Fan, L.-Q., & Heck, A. (1998). Impaired Male Sexual Development in Perinatal Sprague-Dawley and Long-Evans Hooded Rats Exposed in Utero and Lactationally to p,p’-DDE. In TOX1COLOGICAL SCIENCES (Vol. 45). https://academic.oup.com/toxsci/article/45/2/162/1653877