AOP ID and Title:
Graphical Representation
Status
| Author status | OECD status | OECD project | SAAOP status |
|---|---|---|---|
| Open for comment. Do not cite | Under Review | 1.58 | Included in OECD Work Plan |
Abstract
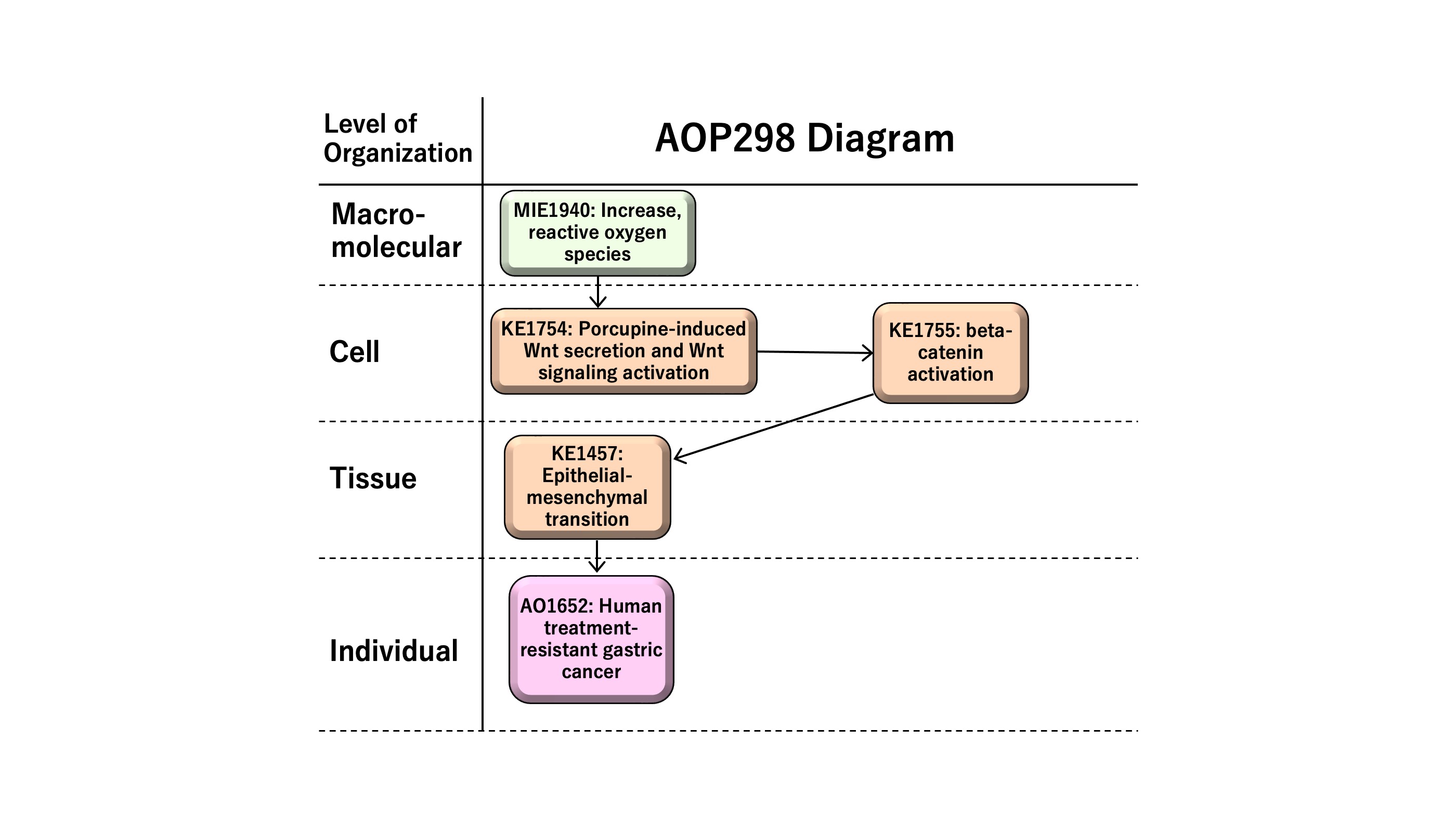
This AOP entitled “Increase in reactive oxygen species (ROS) leading to human treatment-resistant gastric cancer” consists of MIE as KE1115 “Increase, ROS” followed by KE1 as KE1753 “Chronic ROS,” KE2 as KE1754 “porcupine-induced Wnt secretion and Wnt signaling activation,” KE3 as KE1755 “beta-catenin activation,” KE4 as KE1457 “epithelial-mesenchymal transition (EMT),” and AO as KE1651 “human treatment-resistant gastric cancer.” ROS has multiple roles in disease, such as the development and progression of cancer or apoptotic induction, causing anti-tumor effects. In this AOP, we focus on sustained levels of chronic reactive oxygen species (ROS) in inducing therapy resistance in human gastric cancer. Epithelial-mesenchymal transition (EMT), a cellular phenotypic change from epithelial to mesenchymal-like features, demonstrates cancer stem cell-like characteristics in human gastric cancer. EMT is induced by Wnt/beta-catenin signaling, providing the rationale to have Wnt secretion and beta-catenin activation as KE1 and KE2 on the AOP, respectively. The AOP might be useful for the development of anti-cancer drugs or the prediction of adverse effects of therapeutics, which are of possible regulatory relevance.
Summary of the AOP
Events
Molecular Initiating Events (MIE), Key Events (KE), Adverse Outcomes (AO)
| Sequence | Type | Event ID | Title | Short name |
|---|---|---|---|---|
| MIE | 1115 | Increase, Reactive oxygen species | Increase, ROS | |
| 3 | KE | 1754 | Increase, porcupine-induced Wnt secretion and Wnt signaling activation | Increase, porcupine-induced Wnt secretion and Wnt signaling activation |
| 4 | KE | 1755 | beta-catenin activation | beta-catenin activation |
| KE | 1457 | Epithelial Mesenchymal Transition | EMT | |
| 6 | AO | 1651 | Treatment-resistant gastric cancer | Resistant gastric cancer |
Key Event Relationships
| Upstream Event | Relationship Type | Downstream Event | Evidence | Quantitative Understanding |
|---|---|---|---|---|
| Increase, Reactive oxygen species | adjacent | Increase, porcupine-induced Wnt secretion and Wnt signaling activation | Moderate | Moderate |
| Increase, porcupine-induced Wnt secretion and Wnt signaling activation | adjacent | beta-catenin activation | Moderate | Moderate |
| beta-catenin activation | adjacent | Epithelial Mesenchymal Transition | Moderate | Moderate |
| Epithelial Mesenchymal Transition | adjacent | Treatment-resistant gastric cancer | Moderate | Moderate |
Stressors
| Name | Evidence |
|---|---|
| Wnt | High |
| WNT2 | High |
| Porcupine | Moderate |
| Wntless | Moderate |
| Ionizing Radiation | Moderate |
| ferric nitrilotriacetate | Not Specified |
Wnt
WNT induces EMT (J. Zhang, Tian, & Xing, 2016).
WNT2
WNT2 induces EMT in cervical cancer (Zhou et al., 2016).
Porcupine
Porcupine palmitoleates Wnt and facilitates the secretion of the Wnt ligand (Yu & Virshup, 2014) .
Wntless
Wntless binds to and transport Wnt to the plasma membrane leading to the secretion of Wnt ligand (Yu & Virshup, 2014).
ferric nitrilotriacetate
Carcinogenic iron(III)-nitrilotriacetate induces reactive oxygen species production via trasfer of an electron to molecular oxygen to form reactive oxygen species [Tsuchiya K, Akai K, Tokumura A, Abe S, Tamaki T, Takiguchi Y, Fukuzawa K. Biochim Biophys Acta. 2005 Aug 30;1725(1):111-9. doi: 10.1016/j.bbagen.2005.05.001, Akai K, Tsuchiya K, Tokumura A, Kogure K, Ueno S, Shibata A, Tamaki T, Fukuzawa K. Free Radic Res. 2004 Sep;38(9):951-62. doi: 10.1080/1071576042000261945.].
Overall Assessment of the AOP
|
1. Support for Biological Plausibility of KERs |
|
|
MIE => KE1: |
Biological Plausibility of the MIE => KE1 is moderate. |
|
KE1 => KE2: |
Biological Plausibility of the KE1 => KE2 is moderate. |
|
KE2 => KE3: |
Biological Plausibility of the KE2 => KE3 is moderate. EMT-related transcription factors including Snail, ZEB and Twist are up-regulated in cancer cells (Diaz et al., 2014). The transcription factors such as Snail, ZEB and Twist bind to E-cadherin (CDH1) promoter and inhibit the CDH1 transcription via the consensus E-boxes (5’-CACCTG-3’ or 5’-CAGGTG-3’), which leads to EMT (Diaz et al., 2014). |
|
KE3 => AO: |
Biological Plausibility of the KE3 => AO is moderate. EMT phenomenon is related to cancer metastasis and cancer therapy resistance (Smith & Bhowmick, 2016; Tanabe, 2013). Increase expression of enzymes that degrade the extracellular matrix components and the decrease in adhesion to the basement membrane in EMT induce the cell escape from the basement membrane and metastasis (Smith & Bhowmick, 2016). Morphological changes observed during EMT is associated with therapy resistance (Smith & Bhowmick, 2016). |
|
2. Support for essentiality of KEs |
|
|
MIE: Increase, ROS |
Essentiality of the MIE is high. Rationale for Essentiality of the MIE in the AOP: Sustained ROS contributes to the initiation and development of human gastric cancer (Gu et al., 2018). |
|
KE1: Porcupine-induced Wnt secretion and Wnt signaling activation |
Essentiality of the KE1 is moderate. |
|
KE2: beta-catenin activation |
Essentiality of the KE2 is moderate. |
|
KE3: Epithelial-mesenchymal transition (EMT) |
Essentiality of the KE3 is moderate. |
Domain of Applicability
Life Stage Applicability| Life Stage | Evidence |
|---|---|
| All life stages | High |
| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| Homo sapiens | Homo sapiens | High | NCBI |
| Sex | Evidence |
|---|---|
| Unspecific | High |
The AOP298 applies to Homo sapiens (human), all life stages, and both male and female.
Essentiality of the Key Events
Sustained ROS contributes to the initiation and development of human gastric cancer (Gu et al., 2018).
Wnt signaling is involved in cancer malignancy (Tanabe, 2018).
Upon stimulation with Wnt ligand to the Frizzled receptor, Wnt/beta-catenin signaling is activated. Wnt/beta-catenin consists of GSK3 beta inactivation, beta-catenin activation, and up-regulation of transcription factors such as Zeb, Twist, and Snail. The transcription factors Zeb, Twist and Snail relate to the activation of EMT-related genes. EMT is regulated with various gene networks (Tanabe, 2015c, Tanabe et al, 2020a, 2020b).
| Event | Direct Evidence | Indirect Evidence | No experimental evidence |
| MIE: Increase, ROS | *** | * | |
| KE1: Increase, porcupine-induced Wnt secretion and Wnt signaling activation | * | ** | |
| KE2: beta-catenin activation | * | ** | |
| KE3: Epithelial-Mesenchymal Transition | * | ** |
Weight of Evidence Summary
The Wnt signaling promotes EMT and cancer malignancy in colorectal cancer (Lazarova & Bordonaro, 2017). Although the potential pathways other than Wnt signaling exist in EMT induction and the mechanism underlaid cancer malignancy, Wnt signaling is one of the main pathways to induce EMT and cancer malignancy (Polakis, 2012).
| MIE => KE1: Increases in cellular ROS leads to porcupine-induced Wnt secretion and Wnt signaling activation |
Empirical Support of the MIE => KE1 is moderate. Rationale: Production of ROS and DNA double-strand break cause the tissue damages (Gao et al., 2019). ROS-related signaling induces Wnt/b-catenin pathway activation (Pérez et al., 2017). |
| KE1 => KE2: Porcupine-induced Wnt secretion and Wnt signaling activation leads to beta-catenin activation |
Empirical Support of the KE1 => KE2 is moderate. Rationale: Dishevelled (DVL), a positive regulator of Wnt signaling, form the complex with FZD and lead to trigger the Wnt signaling together with Wnt coreceptor low-density lipoprotein (LDL) receptor-related protein 6 (LRP6) (Clevers & Nusse, 2012; Jiang et al., 2015). Wnt binds to FZD and activate the Wnt signaling (Clevers & Nusse, 2012; Janda et al., 2012; Nile et al., 2017). Wnt binding towards FZD induce the formation of the protein complex with LRP5/6 and DVL, leading to the down-stream signaling activation including beta-catenin (Clevers & Nusse, 2012). |
| KE2 => KE3: beta-catenin activation leads to Epithelial-mesenchymal transition (EMT) |
Empirical Support of the KE2 => KE3 is moderate. Rationale: The inhibition of c-MET, which is overexpressed in diffuse-type gastric cancer, induced increase in phosphorylated b-catenin, decrease in b-catenin and Snail (Sohn et al., 2019). The garcinol, that has anti-cancer effect, increases phosphorylated beta-catenin, decreases b-catenin and ZEB1/ZEB2, and inhibit EMT (Ahmad et al., 2012). The inhibition of sortilin by AF38469 (a sortilin inhibitor) or small interference RNA (siRNA) results in decrease in b-catenin and Twist expression in human glioblastoma cells (Yang W. et al., 2019). Histone deacetylase inhibitors effect on EMT-related transcription factors including ZEB, Twist and Snail (Wawruszak et al., 2019). Snail and Zeb induces EMT and suppress E-cadherin (CDH1) (Batlle et al., 2000; Diaz et al., 2014; Peinado et al., 2007). |
| KE3 => AO: Epithelial-mesenchymal transition (EMT) leads to Treatment-resistant gastric cancer |
Empirical Support of the KE3 => AO is moderate. Rationale: EMT activation induces the expression of multiple members of the ATP-binding cassette (ABC) transporter family, which results in doxorubicin resistance (Saxena et al., 2011; Shibue & Weinberg, 2017). TGFb-1 induced EMT results in the acquisition of cancer stem cell (CSC) like properties (Pirozzi et al., 2011; Shibue & Weinberg, 2017). Snail-induced EMT induces the cancer metastasis and resistance to dendritic cell-mediated immunotherapy (Kudo-Saito et al., 2009). Zinc finger E-box-binding homeobox (ZEB1)-induced EMT results in relief of miR-200-mediated repression of programmed cell death 1 ligand (PD-L1) expression, a major inhibitory ligand for the programmed cell death protein (PD-1) immune-checkpoint protein on CD8+ cytotoxic T lymphocyte (CTL), subsequently the CD8+ T cell immunosuppression and metastasis (Chen et al., 2014). |
Quantitative Consideration
Wnt signaling activates the CSCs to promote cancer malignancy (Reya & Clevers, 2005). The responses in KEs related to Wnt signaling, Frizzled activation, GSK3beta inactivation, beta-catenin activation, Snail, Zeb, and Twist activation are dose-dependently related. The quantification of EMT and cancer malignancy would require further investigation.
Considerations for Potential Applications of the AOP (optional)
AOP entitled “Increase in reactive oxygen species (ROS) and chronic ROS leading to human treatment-resistant gastric cancer” might be utilized for the development and risk assessment of anti-cancer drugs. EMT is involved in the acquisition of drug resistance, which is one of the critical features of cancer malignancy. The assessment of the activity of the EMT network would serve as a prediction of the adverse effects of or responsiveness to anti-cancer drugs (Tanabe et al., 2023). The detection methods for increases in ROS in this AOP have future regulatory potentials to assess the human health effects of radiation or ROS-related diseases. The detection methods for human treatment-resistant gastric cancer have future regulatory potentials to diagnose the diseases.
References
Gu, H., Huang, Y. Shen, Y. Liu, F. Zhou, Y. Jin, et al. (2018). Reactive Oxygen Species-Mediated Tumor Microenvironment Transformation: The Mechanism of Radioresistant Gastric Cancer. Oxid Med Cell Longev 2018 Vol. 2018 Pages 5801209. doi:10.1155/2018/5801209
Kudo-Saito, C., Shirako, H., Takeuchi, T., & Kawakami, Y. (2009). Cancer Metastasis Is Accelerated through Immunosuppression during Snail-Induced EMT of Cancer Cells. Cancer Cell, 15(3), 195-206. doi: 10.1016/j.ccr.2009.01.023
Nile, A. H., Mukund, S., Stanger, K., Wang, W., & Hannoush, R. N. (2017). Unsaturated fatty acyl recognition by Frizzled receptors mediates dimerization upon Wnt ligand binding. Proc Natl Acad Sci U S A, 114(16), 4147-4152. doi:10.1073/pnas.1618293114
Tanabe, S. (2018). Wnt Signaling and Epithelial-Mesenchymal Transition Network in Cancer. Research Journal of Oncology, 2(2), 3.
Tanabe, S., Quader, S., Cabral, H., & Ono, R. (2020a). Interplay of EMT and CSC in Cancer and the Potential Therapeutic Strategies. Front. Pharmacol. 11:904. doi: 10.3389/fphar.2020.00904
Tanabe, S., Quader, S., Ono, R., Cabral, H., Aoyagi, K., Hirose, A., Yokozaki, H., & Sasaki, H. (2020b). Molecular Network Profiling in Intestinal- and Diffuse-Type Gastric Cancer. Cancers, 12(12), 3833. https://doi.org/10.3390/cancers12123833
Tanabe, S., Quader, S., Ono, R., Cabral, H., Aoyagi, K., Hirose, A., Perkins, E.J., Yokozaki, H., & Sasaki H. (2023). Regulation of Epithelial–Mesenchymal Transition Pathway and Artificial Intelligence-Based Modeling for Pathway Activity Prediction. Onco, 3(1):13-25. doi: 10.3390/onco3010002
Appendix 1
List of MIEs in this AOP
Event: 1115: Increase, Reactive oxygen species
Short Name: Increase, ROS
Key Event Component
| Process | Object | Action |
|---|---|---|
| reactive oxygen species biosynthetic process | reactive oxygen species | increased |
AOPs Including This Key Event
Biological Context
| Level of Biological Organization |
|---|
| Cellular |
Cell term
| Cell term |
|---|
| cell |
Organ term
| Organ term |
|---|
| organ |
Domain of Applicability
Taxonomic Applicability| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| Vertebrates | Vertebrates | High | NCBI |
| human | Homo sapiens | Moderate | NCBI |
| human and other cells in culture | human and other cells in culture | Moderate | NCBI |
| mouse | Mus musculus | Moderate | NCBI |
| crustaceans | Daphnia magna | High | NCBI |
| Lemna minor | Lemna minor | High | NCBI |
| zebrafish | Danio rerio | High | NCBI |
| Life Stage | Evidence |
|---|---|
| All life stages | High |
| Sex | Evidence |
|---|---|
| Unspecific | High |
| Mixed | High |
ROS is a normal constituent found in all organisms, lifestages, and sexes.
Key Event Description
Biological State: increased reactive oxygen species (ROS)
Biological compartment: an entire cell -- may be cytosolic, may also enter organelles.
Reactive oxygen species (ROS) are O2- derived molecules that can be both free radicals (e.g. superoxide, hydroxyl, peroxyl, alcoxyl) and non-radicals (hypochlorous acid, ozone and singlet oxygen) (Bedard and Krause 2007; Ozcan and Ogun 2015). ROS production occurs naturally in all kinds of tissues inside various cellular compartments, such as mitochondria and peroxisomes (Drew and Leeuwenburgh 2002; Ozcan and Ogun 2015). Furthermore, these molecules have an important function in the regulation of several biological processes – they might act as antimicrobial agents or triggers of animal gamete activation and capacitation (Goud et al. 2008; Parrish 2010; Bisht et al. 2017).
However, in environmental stress situations (exposure to radiation, chemicals, high temperatures) these molecules have its levels drastically increased, and overly interact with macromolecules, namely nucleic acids, proteins, carbohydrates and lipids, causing cell and tissue damage (Brieger et al. 2012; Ozcan and Ogun 2015).
Reactive oxygen species (ROS) refers to the chemical species superoxide, hydrogen peroxide, and their secondary reactive products. In the biological context, ROS are signaling molecules with important roles in cell energy metabolism, cell proliferation, and fate. Therefore, balancing ROS levels at the cellular and tissue level is an important part of many biological processes. Disbalance, mainly an increase in ROS levels, can cause cell dysfunction and irreversible cell damage.
ROS are produced from both exogenous stressors and normal endogenous cellular processes, such as the mitochondrial electron transport chain (ETC). Inhibition of the ETC can result in the accumulation of ROS. Exposure to chemicals, heavy metal ions, or ionizing radiation can also result in increased production of ROS. Chemicals and heavy metal ions can deplete cellular antioxidants reducing the cell’s ability to control cellular ROS and resulting in the accumulation of ROS. Cellular antioxidants include glutathione (GSH), protein sulfhydryl groups, superoxide dismutase (SOD).
ROS are radicals, ions, or molecules that have a single unpaired electron in their outermost shell of electrons, which can be categorized into two groups: free oxygen radicals and non-radical ROS [Liou et al., 2010].
<Free oxygen radicals>
|
superoxide |
O2·- |
|
hydroxyl radical |
·OH |
|
nitric oxide |
NO· |
|
organic radicals |
R· |
|
peroxyl radicals |
ROO· |
|
alkoxyl radicals |
RO· |
|
thiyl radicals |
RS· |
|
sulfonyl radicals |
ROS· |
|
thiyl peroxyl radicals |
RSOO· |
|
disulfides |
RSSR |
<Non-radical ROS>
|
hydrogen peroxide |
H2O2 |
|
singlet oxygen |
1O2 |
|
ozone/trioxygen |
O3 |
|
organic hydroperoxides |
ROOH |
|
hypochlorite |
ClO- |
|
peroxynitrite |
ONOO- |
|
nitrosoperoxycarbonate anion |
O=NOOCO2- |
|
nitrocarbonate anion |
O2NOCO2- |
|
dinitrogen dioxide |
N2O2 |
|
nitronium |
NO2+ |
|
highly reactive lipid- or carbohydrate-derived carbonyl compounds |
|
Potential sources of ROS include NADPH oxidase, xanthine oxidase, mitochondria, nitric oxide synthase, cytochrome P450, lipoxygenase/cyclooxygenase, and monoamine oxidase [Granger et al., 2015]. ROS are generated through NADPH oxidases consisting of p47phox and p67phox. ROS are generated through xanthine oxidase activation in sepsis [Ramos et al., 2018]. Arsenic produces ROS [Zhang et al., 2011]. Mitochondria-targeted paraquat and metformin mediate ROS production [Chowdhury et al., 2020]. ROS are generated by bleomycin [Lu et al., 2010]. Radiation induces dose-dependent ROS production [Ji et al., 2019].
ROS are generated in the course of cellular respiration, metabolism, cell signaling, and inflammation [Dickinson and Chang 2011; Egea et al. 2017]. Hydrogen peroxide is also made by the endoplasmic reticulum in the course of protein folding. Nitric oxide (NO) is produced at the highest levels by nitric oxide synthase in endothelial cells and phagocytes. NO production is one of the main mechanisms by which phagocytes kill bacteria [Wang et al., 2017]. The other species are produced by reactions with superoxide or peroxide, or by other free radicals or enzymes.
ROS activity is principally local. Most ROS have short half-lives, ranging from nano- to milliseconds, so diffusion is limited, while reactive nitrogen species (RNS) nitric oxide or peroxynitrite can survive long enough to diffuse across membranes [Calcerrada et al. 2011]. Consequently, local concentrations of ROS are much higher than average cellular concentrations, and signaling is typically controlled by colocalization with redox buffers [Dickinson and Chang 2011; Egea et al. 2017].
Although their existence is limited temporally and spatially, ROS interact with other ROS or with other nearby molecules to produce more ROS and participate in a feedback loop to amplify the ROS signal, which can increase RNS. Both ROS and RNS also move into neighboring cells, and ROS can increase intracellular ROS signaling in neighboring cells [Egea et al. 2017].
In the primary event, photoreactive chemicals are excited by the absorption of photon energy. The energy of the photoactivated chemicals transfer to oxygen and then generates the reactive oxygen species (ROS), including superoxide (O2−) via type I reaction and singlet oxygen (1O2) via type II reaction, as principal intermediate species in phototoxic reaction (Foote, 1991, Onoue et al. , 2009).
How it is Measured or Detected
Photocolorimetric assays (Sharma et al. 2017; Griendling et al. 2016) or through commercial kits purchased from specialized companies.
Yuan, Yan, et al., (2013) described ROS monitoring by using H2-DCF-DA, a redox-sensitive fluorescent dye. Briefly, the harvested cells were incubated with H2-DCF-DA (50 µmol/L final concentration) for 30 min in the dark at 37°C. After treatment, cells were immediately washed twice, re-suspended in PBS, and analyzed on a BD-FACS Aria flow cytometry. ROS generation was based on fluorescent intensity which was recorded by excitation at 504 nm and emission at 529 nm.
Lipid peroxidation (LPO) can be measured as an indicator of oxidative stress damage Yen, Cheng Chien, et al., (2013).
Chattopadhyay, Sukumar, et al. (2002) assayed the generation of free radicals within the cells and their extracellular release in the medium by addition of yellow NBT salt solution (Park et al., 1968). Extracellular release of ROS converted NBT to a purple colored formazan. The cells were incubated with 100 ml of 1 mg/ml NBT solution for 1 h at 37 °C and the product formed was assayed at 550 nm in an Anthos 2001 plate reader. The observations of the ‘cell-free system’ were confirmed by cytological examination of parallel set of explants stained with chromogenic reactions for NO and ROS.
On the basis of the pathogenesis of drug-induced phototoxicity, a reactive oxygen species (ROS) assay was proposed to evaluate the phototoxic risk of chemicals. The ROS assay can monitor generation of ROS, such as singlet oxygen and superoxide, from photoirradiated chemicals, and the ROS data can be used to evaluate the photoreactivity of chemicals (Onoue et al. , 2014, Onoue et al. , 2013, Onoue and Tsuda, 2006). The ROS assay is a recommended approach by guidelines to evaluate the phototoxic risk of chemicals (ICH, 2014, PCPC, 2014).
<Direct detection>
Many fluorescent compounds can be used to detect ROS, some of which are specific, and others are less specific.
・ROS can be detected by fluorescent probes such as p-methoxy-phenol derivative [Ashoka et al., 2020].
・Chemiluminescence analysis can detect the superoxide, where some probes have a wider range for detecting hydroxyl radical, hydrogen peroxide, and peroxynitrite [Fuloria et al., 2021].
・ROS in the blood can be detected using superparamagnetic iron oxide nanoparticles (SPION)-based biosensor [Lee et al., 2020].
・Hydrogen peroxide (H2O2) can be detected with a colorimetric probe, which reacts with H2O2 in a 1:1 stoichiometry to produce a bright pink colored product, followed by the detection with a standard colorimetric microplate reader with a filter in the 540-570 nm range.
・The levels of ROS can be quantified using multiple-step amperometry using a stainless steel counter electrode and non-leak Ag|AgCl reference node [Flaherty et al., 2017].
・Singlet oxygen can be measured by monitoring the bleaching of p-nitrosodimethylaniline at 440 nm using a spectrophotometer with imidazole as a selective acceptor of singlet oxygen [Onoue et al., 2014].
<Indirect Detection>
Alternative methods involve the detection of redox-dependent changes to cellular constituents such as proteins, DNA, lipids, or glutathione [Dickinson and Chang 2011; Wang et al. 2013; Griendling et al. 2016]. However, these methods cannot generally distinguish between the oxidative species behind the changes and cannot provide good resolution for the kinetics of oxidative activity.
References
Akai, K., et al. (2004). "Ability of ferric nitrilotriacetate complex with three pH-dependent conformations to induce lipid peroxidation." Free Radic Res. Sep;38(9):951-62. doi: 10.1080/1071576042000261945
Ashoka, A. H., et al. (2020). "Recent Advances in Fluorescent Probes for Detection of HOCl and HNO." ACS omega, 5(4), 1730-1742. doi:10.1021/acsomega.9b03420
B.H. Park, S.M. Fikrig, E.M. Smithwick Infection and nitroblue tetrazolium reduction by neutrophils: a diagnostic aid Lancet, 2 (1968), pp. 532-534
Bedard, Karen, and Karl-Heinz Krause. 2007. “The NOX Family of ROS-Generating NADPH Oxidases: Physiology and Pathophysiology.” Physiological Reviews 87 (1): 245–313.
Bisht, Shilpa, Muneeb Faiq, Madhuri Tolahunase, and Rima Dada. 2017. “Oxidative Stress and Male Infertility.” Nature Reviews. Urology 14 (8): 470–85.
Brieger, K., S. Schiavone, F. J. Miller Jr, and K-H Krause. 2012. “Reactive Oxygen Species: From Health to Disease.” Swiss Medical Weekly 142 (August): w13659.
Calcerrada, P., et al. (2011). "Nitric oxide-derived oxidants with a focus on peroxynitrite: molecular targets, cellular responses and therapeutic implications." Curr Pharm Des 17(35): 3905-3932.
Chattopadhyay, Sukumar, et al. "Apoptosis and necrosis in developing brain cells due to arsenic toxicity and protection with antioxidants." Toxicology letters 136.1 (2002): 65-76.
Chowdhury, A. R., et al. (2020). "Mitochondria-targeted paraquat and metformin mediate ROS production to induce multiple pathways of retrograde signaling: A dose-dependent phenomenon." Redox Biol. doi: 10.1016/j.redox.2020.101606. PMID: 32604037; PMCID: PMC7327929.
Dickinson, B. C. and Chang C. J. (2011). "Chemistry and biology of reactive oxygen species in signaling or stress responses." Nature chemical biology 7(8): 504-511.
Drew, Barry, and Christiaan Leeuwenburgh. 2002. “Aging and the Role of Reactive Nitrogen Species.” Annals of the New York Academy of Sciences 959 (April): 66–81.
Egea, J., et al. (2017). "European contribution to the study of ROS: A summary of the findings and prospects for the future from the COST action BM1203 (EU-ROS)." Redox biology 13: 94-162.
Flaherty, R. L., et al. (2017). "Glucocorticoids induce production of reactive oxygen species/reactive nitrogen species and DNA damage through an iNOS mediated pathway in breast cancer." Breast Cancer Research, 19(1), 1–13. https://doi.org/10.1186/s13058-017-0823-8
Foote CS. Definition of type I and type II photosensitized oxidation. Photochem Photobiol. 1991;54:659.
Fuloria, S., et al. (2021). "Comprehensive Review of Methodology to Detect Reactive Oxygen Species (ROS) in Mammalian Species and Establish Its Relationship with Antioxidants and Cancer." Antioxidants (Basel, Switzerland) 10(1) 128. doi:10.3390/antiox10010128
Go, Y. M. and Jones, D. P. (2013). "The redox proteome." J Biol Chem 288(37): 26512-26520.
Goud, Anuradha P., Pravin T. Goud, Michael P. Diamond, Bernard Gonik, and Husam M. Abu-Soud. 2008. “Reactive Oxygen Species and Oocyte Aging: Role of Superoxide, Hydrogen Peroxide, and Hypochlorous Acid.” Free Radical Biology & Medicine 44 (7): 1295–1304.
Granger, D. N. and Kvietys, P. R. (2015). "Reperfusion injury and reactive oxygen species: The evolution of a concept" Redox Biol. doi: 10.1016/j.redox.2015.08.020. PMID: 26484802; PMCID: PMC4625011.
Griendling, K. K., et al. (2016). "Measurement of Reactive Oxygen Species, Reactive Nitrogen Species, and Redox-Dependent Signaling in the Cardiovascular System: A Scientific Statement From the American Heart Association." Circulation research 119(5): e39-75.
Griendling, Kathy K., Rhian M. Touyz, Jay L. Zweier, Sergey Dikalov, William Chilian, Yeong-Renn Chen, David G. Harrison, Aruni Bhatnagar, and American Heart Association Council on Basic Cardiovascular Sciences. 2016. “Measurement of Reactive Oxygen Species, Reactive Nitrogen Species, and Redox-Dependent Signaling in the Cardiovascular System: A Scientific Statement From the American Heart Association.” Circulation Research 119 (5): e39–75.
ICH. ICH Guideline S10 Guidance on Photosafety Evaluation of Pharmaceuticals.: International Council on Harmonisation of Technical Requirements for Registration of Pharmaceuticals for Human Use; 2014.
Itziou, A., et al. (2011). "In vivo and in vitro effects of metals in reactive oxygen species production, protein carbonylation, and DNA damage in land snails Eobania vermiculata." Archives of Environmental Contamination and Toxicology, 60(4), 697–707. https://doi.org/10.1007/s00244-010-9583-5
Ji, W. O., et al. "Quantitation of the ROS production in plasma and radiation treatments of biotargets." Sci Rep. 2019 Dec 27;9(1):19837. doi: 10.1038/s41598-019-56160-0. PMID: 31882663; PMCID: PMC6934759.
Kruk, J. and Aboul-Enein, H. Y. (2017). "Reactive Oxygen and Nitrogen Species in Carcinogenesis: Implications of Oxidative Stress on the Progression and Development of Several Cancer Types." Mini-Reviews in Medicinal Chemistry, 17:11. doi:10.2174/1389557517666170228115324
Lee, D. Y., et al. (2020). "PEGylated Bilirubin-coated Iron Oxide Nanoparticles as a Biosensor for Magnetic Relaxation Switching-based ROS Detection in Whole Blood." Theranostics, 10(5), 1997-2007. doi:10.7150/thno.39662
Li, Z., et al. (2020). "Inhibition of MiR-25 attenuates doxorubicin-induced apoptosis, reactive oxygen species production and DNA damage by targeting pten." International Journal of Medical Sciences, 17(10), 1415–1427. https://doi.org/10.7150/ijms.41980
Liou, G. Y. and Storz, P. "Reactive oxygen species in cancer." Free Radic Res. 2010 May;44(5):479-96. doi:10.3109/10715761003667554. PMID: 20370557; PMCID: PMC3880197.
Lu, Y., et al. (2010). "Phosphatidylinositol-3-kinase/akt regulates bleomycin-induced fibroblast proliferation and collagen production." American journal of respiratory cell and molecular biology, 42(4), 432–441. https://doi.org/10.1165/rcmb.2009-0002OC
Onoue, S., et al. (2013). "Establishment and intra-/inter-laboratory validation of a standard protocol of reactive oxygen species assay for chemical photosafety evaluation." J Appl Toxicol. 33(11):1241-50. doi: 10.1002/jat.2776. Epub 2012 Jun 13. PMID: 22696462.
Onoue S, Hosoi K, Toda T, Takagi H, Osaki N, Matsumoto Y, et al. Intra-/inter-laboratory validation study on reactive oxygen species assay for chemical photosafety evaluation using two different solar simulators. Toxicology in vitro : an international journal published in association with BIBRA. 2014;28:515-23.
Onoue S, Hosoi K, Wakuri S, Iwase Y, Yamamoto T, Matsuoka N, et al. Establishment and intra-/inter-laboratory validation of a standard protocol of reactive oxygen species assay for chemical photosafety evaluation. Journal of applied toxicology : JAT. 2013;33:1241-50.
Onoue S, Kawamura K, Igarashi N, Zhou Y, Fujikawa M, Yamada H, et al. Reactive oxygen species assay-based risk assessment of drug-induced phototoxicity: classification criteria and application to drug candidates. J Pharm Biomed Anal. 2008;47:967-72.
Onoue S, Seto Y, Gandy G, Yamada S. Drug-induced phototoxicity; an early in vitro identification of phototoxic potential of new drug entities in drug discovery and development. Current drug safety. 2009;4:123-36.
Onoue S, Tsuda Y. Analytical studies on the prediction of photosensitive/phototoxic potential of pharmaceutical substances. Pharmaceutical research. 2006;23:156-64.
Ozcan, Ayla, and Metin Ogun. 2015. “Biochemistry of Reactive Oxygen and Nitrogen Species.” In Basic Principles and Clinical Significance of Oxidative Stress, edited by Sivakumar Joghi Thatha Gowder. Rijeka: IntechOpen.
Parrish, A. R. 2010. “2.27 - Hypoxia/Ischemia Signaling.” In Comprehensive Toxicology (Second Edition), edited by Charlene A. McQueen, 529–42. Oxford: Elsevier.
PCPC. PCPC 2014 safety evaluation guidelines; Chapter 7: Evaluation of Photoirritation and Photoallergy potential. Personal Care Products Council; 2014.
Ramos, M. F. P., et al. (2018). "Xanthine oxidase inhibitors and sepsis." Int J Immunopathol Pharmacol. 32:2058738418772210. doi:10.1177/2058738418772210
Ravanat, J. L., et al. (2014). "Radiation-mediated formation of complex damage to DNA: a chemical aspect overview." Br J Radiol 87(1035): 20130715.
Schutzendubel, A. and Polle, A. (2002). "Plant responses to abiotic stresses: heavy metal-induced oxidative stress and protection by mycorrhization." Journal of Experimental Botany, 53(372), 1351–1365. https://doi.org/10.1093/jexbot/53.372.1351
Seto Y, Kato M, Yamada S, Onoue S. Development of micellar reactive oxygen species assay for photosafety evaluation of poorly water-soluble chemicals. Toxicology in vitro : an international journal published in association with BIBRA. 2013;27:1838-46.
Sharma, Gunjan, Nishant Kumar Rana, Priya Singh, Pradeep Dubey, Daya Shankar Pandey, and Biplob Koch. 2017. “p53 Dependent Apoptosis and Cell Cycle Delay Induced by Heteroleptic Complexes in Human Cervical Cancer Cells.” Biomedicine & Pharmacotherapy = Biomedecine & Pharmacotherapie 88 (April): 218–31.
Silva, R., et al. (2019). "Light exposure during growth increases riboflavin production, reactive oxygen species accumulation and DNA damage in Ashbya gossypii riboflavin-overproducing strains." FEMS Yeast Research, 19(1), 1–7. https://doi.org/10.1093/femsyr/foy114
Tsuchiya K, et al. (2005). "Oxygen radicals photo-induced by ferric nitrilotriacetate complex." Biochim Biophys Acta. 1725(1):111-9. doi:10.1016/j.bbagen.2005.05.001
Wang, J., et al. (2017). "Glucocorticoids Suppress Antimicrobial Autophagy and Nitric Oxide Production and Facilitate Mycobacterial Survival in Macrophages." Scientific reports, 7(1), 982. https://doi.org/10.1038/s41598-017-01174-9
Wang, X., et al. (2013). "Imaging ROS signaling in cells and animals." Journal of molecular medicine 91(8): 917-927.
Yen, Cheng Chien, et al. "Inorganic arsenic causes cell apoptosis in mouse cerebrum through an oxidative stress-regulated signaling pathway." Archives of toxicology 85 (2011): 565-575.
Yuan, Yan, et al. "Cadmium-induced apoptosis in primary rat cerebral cortical neurons culture is mediated by a calcium signaling pathway." PloS one 8.5 (2013): e64330.
Zhang, Z., et al. (2011). "Reactive oxygen species mediate arsenic induced cell transformation and tumorigenesis through Wnt/β-catenin pathway in human colorectal adenocarcinoma DLD1 cells. " Toxicology and Applied Pharmacology, 256(2), 114-121. doi:10.1016/j.taap.2011.07.016
List of Key Events in the AOP
Event: 1754: Increase, porcupine-induced Wnt secretion and Wnt signaling activation
Short Name: Increase, porcupine-induced Wnt secretion and Wnt signaling activation
Key Event Component
| Process | Object | Action |
|---|---|---|
| Wnt protein secretion | protein-serine O-palmitoleoyltransferase porcupine | increased |
AOPs Including This Key Event
| AOP ID and Name | Event Type |
|---|---|
| Aop:298 - Increase in reactive oxygen species (ROS) leading to human treatment-resistant gastric cancer | KeyEvent |
Stressors
| Name |
|---|
| Radiation |
Biological Context
| Level of Biological Organization |
|---|
| Cellular |
Cell term
| Cell term |
|---|
| cell |
Organ term
| Organ term |
|---|
| organ |
Domain of Applicability
Taxonomic Applicability| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| Homo sapiens | Homo sapiens | High | NCBI |
| Mus musculus | Mus musculus | High | NCBI |
| Life Stage | Evidence |
|---|---|
| All life stages | Moderate |
| Sex | Evidence |
|---|---|
| Unspecific | High |
Oligomerization of FZD and low-density lipoprotein receptor-related protein 5/6 (LRP5/6) activates Wnt/beta-catenin signaling in Homo sapiens (Hua et al., 2018).
Key Event Description
Porcupine, which is a trans-membrane endoplasmic reticulum O-acyl transferase, is important for the secretion of Wnt ligands(Saha et al., 2016a). WNTs are secreted proteins that contain 22-24 conserved cysteine residues (Foulquier et al., 2018). The WNT molecules consist of molecular families including WNT1, WNT2, WNT2B/WNT13, WNT3, WNT4, WNT5A, WNT5B, WNT6, WNT7A, WNT7B, WNT8A, WNT8B, WNT10B, WNT11, and WNT16. (Clevers & Nusse, 2012; M. Katoh, 2001; Kusserow et al., 2005)
Wnt proteins consist of 350-400 amino acids (Saito-Diaz et al., 2013).
WNT ligands are known to trigger at least three different downstream signaling cascades including canonical WNT/beta-catenin signaling pathway, non-canonical WNT/Ca2+ pathway, and planer cell polarity (PCP) pathway(De, 2011; Lai, Chien, & Moon, 2009; Willert & Nusse, 2012). WNTs bind to Frizzled proteins, which are seven-pass transmembrane receptors with an extracellular N-terminal cysteine-rich domain (Bhanot et al., 1996; Clevers, 2006). Wnt signaling begins with the binding of Wnt ligand towards the Frizzled receptors (Mohammed et al., 2016).
Wnt ligands bind to Frizzled (FZD) receptors which are seven transmembrane-domain protein receptors (Nile, Mukund, Stanger, Wang, & Hannoush, 2017). At least 10 FZD receptors are identified in human cells. FZD receptor is activated by Wnt ligand binding (MacDonald, Tamai, & He, 2009).
How it is Measured or Detected
- Secretion of WNT requires a number of other dedicated factors including the sortin receptor Wntless (WLS), which binds to Wnt and escorts it to the cell surface (Banziger et al., 2006; Ching & Nusse, 2006)
- Wnt signaling is activated by the gene mutations of the signaling components (Ziv et al., 2017).
- Wnt1, Wnt3a, and Wnt5a protein expression are measured by immunoblotting using antibodies for Wnt1, Wnt3a, and Wnt5a, respectively (J. Du et al., 2016; B. Wang et al., 2017).
- WNT2, of which expression is detected by quantitative PCR, immunoblotting, and immunohistochemistry, induces EMT (Zhou et al., 2016).
- Frizzled receptor protein level on the cell surface is measured by flow cytometry with pan-FZD antibody (Jiang et al., 2015; Zeng et al., 2018). DVL protein level is measured by immunoblotting with anti-DVL2 antibodies (Zeng et al., 2018).
- Fzd mRNA level is measured by quantitative reverse transcription-polymerase chain reaction (RT-PCR) (Zeng et al., 2018).
- The up-regulation of WNT ligand expression occurs in Homo sapiens (B. Wang et al., 2017).
- The Wnt genes play an important role in the secretion from cells, glycosylation, and tight association with the cell surface and extracellular matrix in Drosophila melanogaster (Willert & Nusse, 2012).
References
Banziger, C., Soldini, D., Schutt, C., Zipperlen, P., Hausmann, G., & Basler, K. (2006). Wntless, a conserved membrane protein dedicated to the secretion of Wnt proteins from signaling cells. Cell, 125(3), 509-522. doi:10.1016/j.cell.2006.02.049
Bhanot, P., Brink, M., Samos, C. H., Hsieh, J.-C., Wang, Y., Macke, J. P., . . . Nusse, R. (1996). A new member of the frizzled family from Drosophila functions as a Wingless receptor. Nature, 382, 225. doi:10.1038/382225a0
Ching, W., & Nusse, R. (2006). A dedicated Wnt secretion factor. Cell, 125(3), 432-433. doi:10.1016/j.cell.2006.04.018
Clevers, H. (2006). Wnt/beta-catenin signaling in development and disease. Cell, 127(3), 469-480. doi:10.1016/j.cell.2006.10.018
Clevers, H., & Nusse, R. (2012). Wnt/beta-catenin signaling and disease. Cell, 149(6), 1192-1205. doi:10.1016/j.cell.2012.05.012
De, A. (2011). Wnt/Ca2+ signaling pathway: a brief overview. Acta Biochim Biophys Sin (Shanghai), 43(10), 745-756. doi:10.1093/abbs/gmr079
Du, J., Zu, Y., Li, J., Du, S., Xu, Y., Zhang, L., . . . Yang, C. (2016). Extracellular matrix stiffness dictates Wnt expression through integrin pathway. Sci Rep, 6, 20395. doi:10.1038/srep20395
Foulquier, S., Daskalopoulos, E. P., Lluri, G., Hermans, K. C. M., Deb, A., & Blankesteijn, W. M. (2018). WNT Signaling in Cardiac and Vascular Disease. Pharmacol Rev, 70(1), 68-141. doi:10.1124/pr.117.013896
Hua, Y., Yang, Y., Li, Q., He, X., Zhu, W., Wang, J., & Gan, X. (2018). Oligomerization of Frizzled and LRP5/6 protein initiates intracellular signaling for the canonical WNT/beta-catenin pathway. J Biol Chem, 293(51), 19710-19724. doi:10.1074/jbc.RA118.004434
Jiang, X., Charlat, O., Zamponi, R., Yang, Y., & Cong, F. (2015). Dishevelled promotes Wnt receptor degradation through recruitment of ZNRF3/RNF43 E3 ubiquitin ligases. Mol Cell, 58(3), 522-533. doi:10.1016/j.molcel.2015.03.015
Katoh, M. (2001). Molecular cloning and characterization of human WNT3. International journal of oncology, 19(5), 977-982. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/11604997
Kusserow, A., Pang, K., Sturm, C., Hrouda, M., Lentfer, J., Schmidt, H. A., . . . Holstein, T. W. (2005). Unexpected complexity of the Wnt gene family in a sea anemone. Nature, 433(7022), 156-160. doi:10.1038/nature03158
Lai, S. L., Chien, A. J., & Moon, R. T. (2009). Wnt/Fz signaling and the cytoskeleton: potential roles in tumorigenesis. Cell Res, 19(5), 532-545. doi:10.1038/cr.2009.41
MacDonald, B. T., Tamai, K., & He, X. (2009). Wnt/beta-catenin signaling: components, mechanisms, and diseases. Dev Cell, 17(1), 9-26. doi:10.1016/j.devcel.2009.06.016
Mohammed, M. K., Shao, C., Wang, J., Wei, Q., Wang, X., Collier, Z., . . . Lee, M. J. (2016). Wnt/beta-catenin signaling plays an ever-expanding role in stem cell self-renewal, tumorigenesis and cancer chemoresistance. Genes Dis, 3(1), 11-40. doi:10.1016/j.gendis.2015.12.004
Nile, A. H., Mukund, S., Stanger, K., Wang, W., & Hannoush, R. N. (2017). Unsaturated fatty acyl recognition by Frizzled receptors mediates dimerization upon Wnt ligand binding. Proc Natl Acad Sci U S A, 114(16), 4147-4152. doi:10.1073/pnas.1618293114
Saha, S., Aranda, E., Hayakawa, Y., Bhanja, P., Atay, S., Brodin, N. P., . . . Pollard, J. W. (2016). Macrophage-derived extracellular vesicle-packaged WNTs rescue intestinal stem cells and enhance survival after radiation injury. Nature Communications, 7(1), 13096. doi:10.1038/ncomms13096
Saito-Diaz, K., Chen, T. W., Wang, X., Thorne, C. A., Wallace, H. A., Page-McCaw, A., & Lee, E. (2013). The way Wnt works: components and mechanism. Growth Factors, 31(1), 1-31. doi:10.3109/08977194.2012.752737
Wang, B., Tang, Z., Gong, H., Zhu, L., & Liu, X. (2017). Wnt5a promotes epithelial-to-mesenchymal transition and metastasis in non-small-cell lung cancer. Biosci Rep, 37(6). doi:10.1042/BSR20171092
Willert, K., & Nusse, R. (2012). Wnt proteins. Cold Spring Harb Perspect Biol, 4(9), a007864. doi:10.1101/cshperspect.a007864
Zeng, H., Lu, B., Zamponi, R., Yang, Z., Wetzel, K., Loureiro, J., . . . Cong, F. (2018). mTORC1 signaling suppresses Wnt/beta-catenin signaling through DVL-dependent regulation of Wnt receptor FZD level. Proc Natl Acad Sci U S A, 115(44), E10362-E10369. doi:10.1073/pnas.1808575115
Zhou, Y., Huang, Y., Cao, X., Xu, J., Zhang, L., Wang, J., . . . Zheng, M. (2016). WNT2 Promotes Cervical Carcinoma Metastasis and Induction of Epithelial-Mesenchymal Transition. PLoS One, 11(8), e0160414. doi:10.1371/journal.pone.0160414
Ziv, E., Yarmohammadi, H., Boas, F. E., Petre, E. N., Brown, K. T., Solomon, S. B., . . . Erinjeri, J. P. (2017). Gene Signature Associated with Upregulation of the Wnt/beta-Catenin Signaling Pathway Predicts Tumor Response to Transarterial Embolization. J Vasc Interv Radiol, 28(3), 349-355 e341. doi:10.1016/j.jvir.2016.11.004
Event: 1755: beta-catenin activation
Short Name: beta-catenin activation
Key Event Component
| Process | Object | Action |
|---|---|---|
| regulation of beta-catenin-TCF complex assembly | beta-catenin-TCF complex | occurrence |
AOPs Including This Key Event
| AOP ID and Name | Event Type |
|---|---|
| Aop:298 - Increase in reactive oxygen species (ROS) leading to human treatment-resistant gastric cancer | KeyEvent |
Biological Context
| Level of Biological Organization |
|---|
| Cellular |
Cell term
| Cell term |
|---|
| cell |
Organ term
| Organ term |
|---|
| organ |
Domain of Applicability
Taxonomic Applicability| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| Homo sapiens | Homo sapiens | High | NCBI |
| Life Stage | Evidence |
|---|---|
| All life stages | Moderate |
| Sex | Evidence |
|---|---|
| Unspecific | High |
Beta-catenin is stabilized and translocated into nucleus in Homo sapiens (Huang et al., 2019).
Beta-catenin is activated in Homo sapiens (Huang et al., 2019) (Naujok et al., 2014).
Key Event Description
Upon the Wnt signaling activation, beta-catenin is stabilized and activated via inhibition of the phosphorylation by GSK3beta (Huang et al., 2019). Once the beta-catenin is stabilized, it translocates into the nucleus and enhances the expression of target genes of Wnt/beta-catenin signaling pathway (Huang et al., 2019). Beta-catenin activation is related to cancer (Tanabe, 2014).
Dishevelled (DVL), a positive regulator of Wnt signaling, forms the complex with FZD and leads to trigger the Wnt signaling together with Wnt coreceptor low-density lipoprotein (LDL) receptor-related protein 6 (LRP6) (Clevers & Nusse, 2012; Jiang, et al., 2015). DVL, however, has a controversial role to promote Wnt receptor degradation (Jiang et al., 2015). Meanwhile, DVL-dependent regulation of FZD level is involved in mTORC1 signaling suppression via Wnt/beta-catenin signaling (Zeng et al., 2018). The recruitment of Axin to the DVL-FZD complex induces the beta-catenin stabilization and activation. The stabilized beta-catenin translocates into the nucleus, which forms the complex with TCF to induce the up-regulated expression of proliferation-related genes.
How it is Measured or Detected
The beta-catenin level in nucleus is measured by immunoblotting with anti-beta-catenin antibody (Huang et al., 2019).
The beta-catenin nuclear translocation is measured by immunofluorescence assay (Huang et al., 2019).
Activity of beta-catenin is measured by Wnt/beta-catenin activity assay, in which the vector containing the firefly luciferase gene controlled by TCF/LEF binding sites is transfected in the cells (Naujok et al., 2014).
References
Clevers, H., & Nusse, R. (2012). Wnt/beta-catenin signaling and disease. Cell, 149(6), 1192-1205. doi:10.1016/j.cell.2012.05.012
Huang, J. Q., Wei, F. K., Xu, X. L., Ye, S. X., Song, J. W., Ding, P. K., . . . Gong, L. Y. (2019). SOX9 drives the epithelial-mesenchymal transition in non-small-cell lung cancer through the Wnt/beta-catenin pathway. J Transl Med, 17(1), 143. doi:10.1186/s12967-019-1895-2
Jiang, X., Charlat, O., Zamponi, R., Yang, Y., & Cong, F. (2015). Dishevelled promotes Wnt receptor degradation through recruitment of ZNRF3/RNF43 E3 ubiquitin ligases. Mol Cell, 58(3), 522-533. doi:10.1016/j.molcel.2015.03.015
Naujok, O., Lentes, J., Diekmann, U., Davenport, C., & Lenzen, S. (2014). Cytotoxicity and activation of the Wnt/beta-catenin pathway in mouse embryonic stem cells treated with four GSK3 inhibitors. BMC Res Notes, 7, 273. doi:10.1186/1756-0500-7-273
Tanabe, S. (2014). Role of mesenchymal stem cells in cell life and their signaling. World journal of stem cells, 6(1), 24-32. doi:10.4252/wjsc.v6.i1.24
Zeng, H., Lu, B., Zamponi, R., Yang, Z., Wetzel, K., Loureiro, J., . . . Cong, F. (2018). mTORC1 signaling suppresses Wnt/beta-catenin signaling through DVL-dependent regulation of Wnt receptor FZD level. Proc Natl Acad Sci U S A, 115(44), E10362-E10369. doi:10.1073/pnas.1808575115
Event: 1457: Epithelial Mesenchymal Transition
Short Name: EMT
Key Event Component
| Process | Object | Action |
|---|---|---|
| epithelial to mesenchymal transition | Epithelial cell | occurrence |
AOPs Including This Key Event
Biological Context
| Level of Biological Organization |
|---|
| Cellular |
Cell term
| Cell term |
|---|
| epithelial cell |
Organ term
| Organ term |
|---|
| organ |
Domain of Applicability
Taxonomic Applicability| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| humans | Homo sapiens | High | NCBI |
| Life Stage | Evidence |
|---|---|
| Not Otherwise Specified | Not Specified |
| Sex | Evidence |
|---|---|
| Unspecific | Not Specified |
The key event is applicaple in Homo sapiens:
- Wnt5a expression leads to epithelial-mesenchymal transition (EMT) and metastasis in non-small-cell lung cancer in Homo sapiens (Wang et al., 2017).
- WNT2 expression lead to EMT induction in Homo sapiens (Zhou et al., 2016).
- EMT is induced in cancer and involved in cancer metastasis in Homo sapiens (Suarez-Carmona, Lesage, Cataldo, & Gilles, 2017) (Du & Shim, 2016).
Regulation of miRNA expression by DNA replication,damage and repair responses,transcription and translation has been proved in animals like mice,canine and cell line experiments.
Key Event Description
Epithelial-mesenchymal transition (EMT) is a phenomenon in which the cells transit from epithelial-like into mesenchymal-like phenotypes (Huan et al., 2022; Tanabe, 2017; Tanabe et al., 2015). In cancer, cells exhibiting EMT features contribute to metastasis and drug resistance.
It is known that D-2-hydroxyglurate induces EMT (Guerra et al., 2017; Jia et al., 2018; Mishra et al., 2018; Sciacovelli & Frezza, 2017). D-2-hydroxyglurate, an inhibitor of Jumonji-family histone demethylase, increased the trimethylation of histone H3 lysine 4 (H3K4) in the promoter region of the zinc finger E-box-binding homeobox 1 (ZEB1), followed by the induction of EMT (Colvin et al., 2016).
Wnt5a induces EMT and metastasis in non-small-cell lung cancer (Wang et al., 2017).
EMT is related to Wnt/beta-catenin signaling and is important for treatment-resistant cancer (Tanabe et al., 2016).
TGFbeta induces EMT (Wendt et al., 2010).
ZEB is one of the critical transcription factors for EMT regulation (Zhang et al., 2015).
SNAI1 (Snail) is an important transcription factor for cell differentiation and survival. The phosphorylation and nuclear localization of Snail1 induced by Wnt signaling pathways are critical for the regulation of EMT (Kaufhold & Bonavida, 2014).
Transcription factors SNAI1 and TWIST1 induce EMT (Hodge et al., 2018) (Mani et al., 2008).
It is suggested that Sp1, a transcription factor involved in cell growth and metastasis, is induced by cytochrome P450 1B1 (CYP1B1), and promotes EMT, which leads to cell proliferation and metastasis (Kwon et al., 2016).
|
Biological state
An epithelial-mesenchymal transition (EMT) is a biologic process in which epithelial cells are polarized, interact through their basal surface with basement membrane, and undergo biochemical changes to assume a mesenchymal cell phenotype. This phenotypic transformation has various characters such as enhanced migratory capacity, high invasiveness, elevated resistance to apoptosis, and greatly increased production of ECM components (Kalluri, R., and Neilson, E.G. 2003). The completion of an EMT is signalled by the degradation of the underlying basement membrane and the formation of a mesenchymal cell that can migrate away from the epithelial layer in which it originated.
EMT has a number of distinct molecular processes like activation of transcription factors, expression of specific cell surface proteins, reorganization and expression of cytoskeletal proteins, production of ECM-degrading enzymes, and changes in the expression of specific microRNAs. These factors are used as biomarkers to demonstrate the passage of a cell through an EMT.
Biological compartment Cellular Role in General Biology: Excessive proliferation of epithelial cells and angiogenesis mark the initiation and early growth of primary epithelial cancers. (Hanahan, D., and Weinberg, R.A. 2000). The subsequent acquisition of invasiveness, initially manifest by invasion through the basement membrane, is thought to herald the onset of the last stages of the multi-step process that leads eventually to metastatic dissemination, with life-threatening consequences. There has been an intense research going on in the genetic controls and biochemical mechanisms underlying the acquisition of the invasive phenotype and the subsequent systemic spread of the cancer cell. Activation of an EMT program has been proposed as the critical mechanism for the acquisition of malignant phenotypes by epithelial cancer cells (Thiery, J.P. 2002). Pre-clinical experiments such as mice models and cell culture experiments has demonstrated that carcinoma cells can acquire a mesenchymal phenotype and express mesenchymal markers such as α-SMA, FSP1, vimentin, and desmin (Yang, J., and Weinberg, R.A. 2008). These cells are seen at the invasive front of primary tumors and are considered to be the cells that eventually enter into subsequent steps of the invasion-metastasis cascade, i.e., intravasation, transport through the circulation, extravasation, formation of micro metastases, and ultimately colonization (the growth of small colonies into macroscopic metastases) (Thiery, J.P. 2002, Fidler, I.J., and Poste, G. 2008, Brabletz, T., et al. 2001). An apparent paradox comes from the observation that the EMT-derived migratory cancer cells typically establish secondary colonies at distant sites that resemble, at the histopathological level, the primary tumor from which they arose; accordingly, they no longer exhibit the mesenchymal phenotypes ascribed to metastasizing carcinoma cells. Reconciling this behaviour with the proposed role of EMT as a facilitator of metastatic dissemination requires the additional notion that metastasizing cancer cells must shed their mesenchymal phenotype via a MET during the course of secondary tumor formation (Zeisberg, M et al 2005). The tendency of disseminated cancer cells to undergo EMT likely reflects the local microenvironments that they encounter after extravasation into the parenchyma of a distant organ, quite possibly the absence of the heterotypic signals they experienced in the primary tumor that were responsible for inducing the EMT in the first place (Thiery, J.P. 2002, Jechlinger, M et al 2002, Bissell, M.J et al 2002). These evidences indicate that induction of an EMT is likely to be a centrally important mechanism for the progression of carcinomas to a metastatic stage and implicates MET during the subsequent colonization process. However, many steps of this mechanistic model still require direct experimental validation. It remains unclear at present whether these phenomena and molecular mechanisms relate to and explain the metastatic dissemination of non-epithelial cancer cells. The entire spectrum of signaling agents that contribute to EMTs of carcinoma cells remains unclear. One theory suggests that the genetic and epigenetic alterations undergone by cancer cells during the course of primary tumor formation render them especially responsive to EMT-inducing heterotypic signals originating in the tumor-associated stroma. Oncogenes induce senescence, and recent studies suggest that cancer cell EMTs may also play a role in preventing senescence induced by oncogenes, thereby facilitating subsequent aggressive dissemination (Smit, M.A., and Peeper, D.S. 2008, Ansieau, S., et al. 2008, Weinberg, R.A. 2008). In the case of many carcinomas, EMT-inducing signals emanating from the tumor-associated stroma, notably HGF, EGF, PDGF, and TGF-β, appear to be responsible for the induction or functional activation in cancer cells of a series of EMT-inducing transcription factors, notably Snail, Slug, zinc finger E-box binding homeobox 1 (ZEB1), Twist, Goosecoid, and FOXC2 (Thiery, J.P. 2002, Jechlinger, M et al 2002, Shi, Y., and Massague, J. 2003, Niessen, K., et al. 2008, Medici, D et al 2008, Kokudo, T., et al. 2008). Once expressed and activated, each of these transcription factors can act pleiotropically to choreograph the complex EMT program, more often than not with the help of other members of this cohort of transcription factors. The actual implementation by these cells of their EMT program depends on a series of intracellular signaling networks involving, among other signal- transducing proteins, ERK, MAPK, PI3K, Akt, Smads, RhoB, β-catenin, lymphoid enhancer binding factor (LEF), Ras, and c-Fos as well as cell surface proteins such as β4 integrins, α5β1 integrin, and αVβ6 integrin (Tse, J.C., and Kalluri, R. 2007). Activation of EMT programs is also facilitated by the disruption of cell-cell adherens junctions and the cell-ECM adhesions mediated by integrins (Yang, J., and Weinberg, R.A. 2008, Weinberg, R.A. 2008, Gupta, P.B et al 2005, Yang, J et al 2006, Mani, S.A., et al. 2007, Mani, S.A., et al. 2008, Hartwell, K.A., et al. 2006, Taki, M et al 2006)..
|
How it is Measured or Detected
Loss of E-cadherin and cell polarity is considered to be a fundamental event in epithelial-mesenchymal transition. The simultaneous expression of epithelial (e.g. E-cadherin) and mesenchymal markers (e.g. N-cadherin and vimentin) within the airway epithelium are indicative for ongoing transition (Borthwick et al. 2009, 2010).
|
|
Method/ measurement referenc |
Reliability
|
Strength of evidence
|
Assay fit for purpose
|
Repeatability/ reproducibility
|
Direct measure |
|
Human cell line |
qRT-PCR,cell viability assay, Western blotting,EdU incorporation assay |
+ |
Strong |
Yes |
Yes |
Yes |
|
Human |
IHC,micro array,qPCR, SNP array |
+ |
Moderate |
Yes |
Yes |
Yes |
- EMT can be detected by immunostaining with pro-surfactant protein-C (pro-SPC) and N-cadherin in idiopathic pulmonary fibrosis (IPF) lung in vivo (Kim et al., 2006).
- EMT can be detected by immunostaining with vimentin in lung alveola in vivo (Kim et al., 2006).
- EMT can be detected as the increased level of the transcription factors, zinc finger E-box-binding homeobox (ZEB), Twist and Snail (Huang et al., 2022).
References
Borthwick, L. A., Parker, S. M., Brougham, K. A., Johnson, G. E., Gorowiec, M. R., Ward, C., … Fisher, A. J. (2009). Epithelial to mesenchymal transition (EMT) and airway remodelling after human lung transplantation. Thorax, 64(9), 770–777. https://doi.org/10.1136/thx.2008.104133
Borthwick, L. A., McIlroy, E. I., Gorowiec, M. R., Brodlie, M., Johnson, G. E., Ward, C., … Fisher, A. J. (2010). Inflammation and epithelial to mesenchymal transition in lung transplant recipients: Role in dysregulated epithelial wound repair. American Journal of Transplantation, 10(3), 498–509. https://doi.org/10.1111/j.1600-6143.2009.02953.x
Al Saleh, S., Al Mulla, F., & Luqmani, Y. A. (2011). Estrogen receptor silencing induces epithelial to mesenchymal transition in human breast cancer cells. PloS one, 6(6), e20610.
Bissell, M. J., Radisky, D. C., Rizki, A., Weaver, V. M., & Petersen, O. W. (2002). The organizing principle: microenvironmental influences in the normal and malignant breast. Differentiation, 70(9-10), 537-546.
Bouris, P., Skandalis, S. S., Piperigkou, Z., Afratis, N., Karamanou, K., Aletras, A. J., ... & Karamanos, N. K. (2015). Estrogen receptor alpha mediates epithelial to mesenchymal transition, expression of specific matrix effectors and functional properties of breast cancer cells. Matrix Biology, 43, 42-60.
Brabletz, T., Jung, A., Reu, S., Porzner, M., Hlubek, F., Kunz-Schughart, L. A., ... & Kirchner, T. (2001). Variable β-catenin expression in colorectal cancers indicates tumor progression driven by the tumor environment. Proceedings of the National Academy of Sciences, 98(18), 10356-10361.
Brabletz, T., Jung, A., Reu, S., Porzner, M., Hlubek, F., Kunz-Schughart, L. A., ... & Kirchner, T. (2001). Variable β-catenin expression in colorectal cancers indicates tumor progression driven by the tumor environment. Proceedings of the National Academy of Sciences, 98(18), 10356-10361.
Colvin, H., Nishida, N., Konno, M., Haraguchi, N., Takahashi, H., Nishimura, J., . . . Ishii, H. (2016). Oncometabolite D-2-Hydroxyglurate Directly Induces Epithelial-Mesenchymal Transition and is Associated with Distant Metastasis in Colorectal Cancer. Sci Rep, 6, 36289. doi:10.1038/srep36289
Du, B., & Shim, J. S. (2016). Targeting Epithelial-Mesenchymal Transition (EMT) to Overcome Drug Resistance in Cancer. Molecules, 21(7). doi:10.3390/molecules21070965
Fang, C. X., Ma, C. M., Jiang, L., Wang, X. M., Zhang, N., Ma, J. N., . . . Zhao, Y. D. (2018). p38 MAPK is Crucial for Wnt1- and LiCl-Induced Epithelial Mesenchymal Transition. Curr Med Sci, 38(3), 473-481. doi:10.1007/s11596-018-1903-4
Fidler, I. J., & Poste, G. (2008). The “seed and soil” hypothesis revisited. The lancet oncology, 9(8), 808.
Guerra, F., Guaragnella, N., Arbini, A. A., Bucci, C., Giannattasio, S., & Moro, L. (2017). Mitochondrial Dysfunction: A Novel Potential Driver of Epithelial-to-Mesenchymal Transition in Cancer. Front Oncol, 7, 295. doi:10.3389/fonc.2017.00295
Gupta, P. B., Mani, S., Yang, J., Hartwell, K., & Weinberg, R. A. (2005, January). The evolving portrait of cancer metastasis. In Cold Spring Harbor symposia on quantitative biology (Vol. 70, pp. 291-297). Cold Spring Harbor Laboratory Press.
Hanahan, D., and Weinberg, R.A. (2000). The hall- marks of cancer. Cell. 100:57–70.
Hartwell, K. A., Muir, B., Reinhardt, F., Carpenter, A. E., Sgroi, D. C., & Weinberg, R. A. (2006). The Spemann organizer gene, Goosecoid, promotes tumor metastasis. Proceedings of the National Academy of Sciences, 103(50), 18969-18974.
Jechlinger, M., Grünert, S., & Beug, H. (2002). Mechanisms in epithelial plasticity and metastasis: insights from 3D cultures and expression profiling. Journal of mammary gland biology and neoplasia, 7(4), 415-432.
Hodge, D. Q., Cui, J., Gamble, M. J., & Guo, W. (2018). Histone Variant MacroH2A1 Plays an Isoform-Specific Role in Suppressing Epithelial-Mesenchymal Transition. Sci Rep, 8(1), 841. doi:10.1038/s41598-018-19364-4
Huan, Z., Zhang, Z., Zhou, C., Liu, L., Huang, C. (2022). Epithelial-mesenchymal transition: The history, regulatory mechanism, and cancer therapeutic opportunities. MedComm. 2022 May 18;3(2):e144. doi: 10.1002/mco2.144
Jia, D., Park, J. H., Jung, K. H., Levine, H., & Kaipparettu, B. A. (2018). Elucidating the Metabolic Plasticity of Cancer: Mitochondrial Reprogramming and Hybrid Metabolic States. Cells, 7(3). doi:10.3390/cells7030021
Kalluri, R., & Neilson, E. G. (2003). Epithelial-mesenchymal transition and its implications for fibrosis. The Journal of clinical investigation, 112(12), 1776-1784.
Kaufhold, S., & Bonavida, B. (2014). Central role of Snail1 in the regulation of EMT and resistance in cancer: a target for therapeutic intervention. J Exp Clin Cancer Res, 33, 62. doi:10.1186/s13046-014-0062-0
Kim, K. K., Kugler, M. C., Wolters, P. J., Robillard, L., Galvez, M. G., Brumwell, A. N., . . . Chapman, H. A. (2006). Alveolar epithelial cell mesenchymal transition develops in vivo during pulmonary fibrosis and is regulated by the extracellular matrix. PNAS, 103(35), 13180-13185. doi:10.1073/pnas.0605669103
Kwon, Y. J., Baek, H. S., Ye, D. J., Shin, S., Kim, D., & Chun, Y. J. (2016). CYP1B1 Enhances Cell Proliferation and Metastasis through Induction of EMT and Activation of Wnt/beta-Catenin Signaling via Sp1 Upregulation. PLoS One, 11(3), e0151598. doi:10.1371/journal.pone.0151598
Kokudo, T., Suzuki, Y., Yoshimatsu, Y., Yamazaki, T., Watabe, T., & Miyazono, K. (2008). Snail is required for TGFβ-induced endothelial-mesenchymal transition of embryonic stem cell-derived endothelial cells. Journal of cell science, 121(20), 3317-3324.
Lin, H. Y., Liang, Y. K., Dou, X. W., Chen, C. F., Wei, X. L., Zeng, D., ... & Zhang, G. J. (2018). Notch3 inhibits epithelial–mesenchymal transition in breast cancer via a novel mechanism, upregulation of GATA-3 expression. Oncogenesis, 7(8), 1-15.
Liu, Y., Liu, R., Fu, P., Du, F., Hong, Y., Yao, M., ... & Zheng, S. (2015). N1-Guanyl-1, 7-diaminoheptane sensitizes estrogen receptor negative breast cancer cells to doxorubicin by preventing epithelial-mesenchymal transition through inhibition of eukaryotic translation initiation factor 5A2 activation. Cellular Physiology and Biochemistry, 36(6), 2494-2503.
Mani, S. A., Yang, J., Brooks, M., Schwaninger, G., Zhou, A., Miura, N., ... & Weinberg, R. A. (2007). Mesenchyme Forkhead 1 (FOXC2) plays a key role in metastasis and is associated with aggressive basal-like breast cancers. Proceedings of the National Academy of Sciences, 104(24), 10069-10074.
Mani, S. A., Guo, W., Liao, M. J., Eaton, E. N., Ayyanan, A., Zhou, A. Y., ... & Weinberg, R. A. (2008). The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell, 133(4), 704-715.
Medici, D., Hay, E. D., & Olsen, B. R. (2008). Snail and Slug promote epithelial-mesenchymal transition through β-catenin–T-cell factor-4-dependent expression of transforming growth factor-β3. Molecular biology of the cell, 19(11), 4875-4887.
Mishra, P., Tang, W., Putluri, V., Dorsey, T. H., Jin, F., Wang, F., . . . Ambs, S. (2018). ADHFE1 is a breast cancer oncogene and induces metabolic reprogramming. J Clin Invest, 128(1), 323-340. doi:10.1172/JCI93815
Niessen, K., Fu, Y., Chang, L., Hoodless, P. A., McFadden, D., & Karsan, A. (2008). Slug is a direct Notch target required for initiation of cardiac cushion cellularization. The Journal of cell biology, 182(2), 315-325.
Sciacovelli, M., & Frezza, C. (2017). Metabolic reprogramming and epithelial-to-mesenchymal transition in cancer. FEBS J, 284(19), 3132-3144. doi:10.1111/febs.14090
Shi, Y., & Massagué, J. (2003). Mechanisms of TGF-β signaling from cell membrane to the nucleus. cell, 113(6), 685-700.
Smit, M. A., & Peeper, D. S. (2008). Deregulating EMT and senescence: double impact by a single twist. Cancer cell, 14(1), 5-7.
Suarez-Carmona, M., Lesage, J., Cataldo, D., & Gilles, C. (2017). EMT and inflammation: inseparable actors of cancer progression. Mol Oncol, 11(7), 805-823. doi:10.1002/1878-0261.12095
Sun, J., Yang, X., Zhang, R., Liu, S., Gan, X., Xi, X., . . . Sun, Y. (2017). GOLPH3 induces epithelial-mesenchymal transition via Wnt/beta-catenin signaling pathway in epithelial ovarian cancer. Cancer Med, 6(4), 834-844. doi:10.1002/cam4.1040
Taki, M., Verschueren, K., Yokoyama, K., Nagayama, M., & Kamata, N. (2006). Involvement of Ets-1 transcription factor in inducing matrix metalloproteinase-2 expression by epithelial-mesenchymal transition in human squamous carcinoma cells. International journal of oncology, 28(2), 487-496.
Tanabe, S. (2017). Molecular markers and networks for cancer and stem cells. J Embryol Stem Cell Res, 1(1).
Tanabe, S., Kawabata, T., Aoyagi, K., Yokozaki, H., & Sasaki, H. (2016). Gene expression and pathway analysis of CTNNB1 in cancer and stem cells. World J Stem Cells, 8(11), 384-395. doi:10.4252/wjsc.v8.i11.384
Tanabe, S., Komatsu, M., Kazuhiko, A., Yokozaki, H., & Sasaki, H. (2015). Implications of epithelial-mesenchymal transition in gastric cancer. Translational Gastrointestinal Cancer, 4(4), 258-264
Thiery, J. P. (2002). Epithelial–mesenchymal transitions in tumour progression. Nature reviews cancer, 2(6), 442-454.
Tse, J. C., & Kalluri, R. (2007). Mechanisms of metastasis: epithelial‐to‐mesenchymal transition and contribution of tumor microenvironment. Journal of cellular biochemistry, 101(4), 816-829.
Wang, B., Tang, Z., Gong, H., Zhu, L., & Liu, X. (2017). Wnt5a promotes epithelial-to-mesenchymal transition and metastasis in non-small-cell lung cancer. Biosci Rep, 37(6). doi:10.1042/BSR20171092
Weinberg, R. A. (2008). Twisted epithelial–mesenchymal transition blocks senescence. Nature cell biology, 10(9), 1021-1023.
Wendt, M. K., Smith, J. A., & Schiemann, W. P. (2010). Transforming growth factor-beta-induced epithelial-mesenchymal transition facilitates epidermal growth factor-dependent breast cancer progression. Oncogene, 29(49), 6485-6498. doi:10.1038/onc.2010.377
Wik, E., Ræder, M. B., Krakstad, C., Trovik, J., Birkeland, E., Hoivik, E. A., ... & Salvesen, H. B. (2013). Lack of estrogen receptor-α is associated with epithelial–mesenchymal transition and PI3K alterations in endometrial carcinoma. Clinical Cancer Research, 19(5), 1094-1105.
Yang, J., & Weinberg, R. A. (2008). Epithelial-mesenchymal transition: at the crossroads of development and tumor metastasis. Developmental cell, 14(6), 818-829.
Yang, J., Mani, S. A., & Weinberg, R. A. (2006). Exploring a new twist on tumor metastasis. Cancer research, 66(9), 4549-4552.
Ye, Y., Xiao, Y., Wang, W., Yearsley, K., Gao, J. X., Shetuni, B., & Barsky, S. H. (2010). ERα signaling through slug regulates E-cadherin and EMT. Oncogene, 29(10), 1451-1462.
Zeisberg, M., Shah, A. A., & Kalluri, R. (2005). Bone morphogenic protein-7 induces mesenchymal to epithelial transition in adult renal fibroblasts and facilitates regeneration of injured kidney. Journal of Biological Chemistry, 280(9), 8094-8100.
Zeng, Q., Zhang, P., Wu, Z., Xue, P., Lu, D., Ye, Z., ... & Yan, X. (2014). Quantitative proteomics reveals ER-α involvement in CD146-induced epithelial-mesenchymal transition in breast cancer cells. Journal of proteomics, 103, 153-169.
Zhang, P., Sun, Y., & Ma, L. (2015). ZEB1: at the crossroads of epithelial-mesenchymal transition, metastasis and therapy resistance. Cell Cycle, 14(4), 481-487. doi:10.1080/15384101.2015.1006048
Zhou, Y., Huang, Y., Cao, X., Xu, J., Zhang, L., Wang, J., . . . Zheng, M. (2016). WNT2 Promotes Cervical Carcinoma Metastasis and Induction of Epithelial-Mesenchymal Transition. PLoS One, 11(8), e0160414. doi:10.1371/journal.pone.0160414
List of Adverse Outcomes in this AOP
Event: 1651: Treatment-resistant gastric cancer
Short Name: Resistant gastric cancer
Key Event Component
| Process | Object | Action |
|---|---|---|
| regulation of cellular response to drug | occurrence |
AOPs Including This Key Event
| AOP ID and Name | Event Type |
|---|---|
| Aop:298 - Increase in reactive oxygen species (ROS) leading to human treatment-resistant gastric cancer | AdverseOutcome |
Biological Context
| Level of Biological Organization |
|---|
| Tissue |
Organ term
| Organ term |
|---|
| organ |
Domain of Applicability
Taxonomic Applicability| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| Homo sapiens | Homo sapiens | High | NCBI |
| Life Stage | Evidence |
|---|---|
| All life stages | High |
| Sex | Evidence |
|---|---|
| Unspecific | High |
Drug resistance occurs in Homo sapiens (Du & Shim, 2016).
Key Event Description
It is known that diffuse-type gastric cancer, which has a poor prognosis, is treatment-resistant and more malignant compared to intestinal-type gastric cancer (Tanabe et al., 2014). Drug resistance is involved in EMT, which is an important phenomenon exhibiting features similar to cancer stem cells (CSCs) (Du & Shim, 2016).
EMT is involved in metastasis and cancer therapy resistance (Smith & Bhowmick, 2016).
How it is Measured or Detected
Treatment-resistant gastric cancer and EMT can be detected with biomarkers (Zeisberg & Neilson, 2009).
Treatment-resistant gastric cancer which exhibits EMT phenotype can be detected as the increased level of the transcription factors, zinc finger E-box-binding homeobox 1/2 (ZEB1/2), SNAI1/2, and TWIST2 which are associated with the activation of EMT-related genes (Tanabe et al., 2022a and 2022b).
Regulatory Significance of the AO
Drug resistance is very important in cancer treatment since cancer metastasis and recurrence are some of the main obstacles to treating cancer. Cancer stem cells that share the phenotype of EMT may be targeted in anti-cancer drug development.
References
Du, B., & Shim, J. S. (2016). Targeting Epithelial-Mesenchymal Transition (EMT) to Overcome Drug Resistance in Cancer. Molecules, 21(7). doi:10.3390/molecules21070965
Smith, B. N., & Bhowmick, N. A. (2016). Role of EMT in Metastasis and Therapy Resistance. J Clin Med, 5(2). doi:10.3390/jcm5020017
Tanabe, S., Aoyagi, K., Yokozaki, H., Sasaki, H. (2014). Gene expression signatures for identifying diffuse-type gastric cancer associated with epithelial-mesenchymal transition. International journal of oncology, 44(6), 1955-1970. doi:10.3892/ijo.2014.2387
Tanabe, S., Quader, S., Cabral, H., Ono, R. (2020a). Interplay of EMT and CSC in Cancer and the Potential Therapeutic Strategies. Front Pharmacol, 11, 904. doi:10.3389/fphar.2020.00904
Tanabe S, Quader S, Ono R, Cabral H, Aoyagi K, Hirose A, Yokozaki H., Sasaki, H. (2020b). Molecular Network Profiling in Intestinal- and Diffuse-Type Gastric Cancer. Cancers (Basel), 12(12), 3833. doi:10.3390/cancers12123833
Zeisberg, M., & Neilson, E. G. (2009). Biomarkers for epithelial-mesenchymal transitions. J Clin Invest, 119(6), 1429-1437. doi:10.1172/JCI36183
Appendix 2
List of Key Event Relationships in the AOP
List of Adjacent Key Event Relationships
Relationship: 3505: Increase, ROS leads to Increase, porcupine-induced Wnt secretion and Wnt signaling activation
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Increase in reactive oxygen species (ROS) leading to human treatment-resistant gastric cancer | adjacent | Moderate | Moderate |
Evidence Supporting Applicability of this Relationship
| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| Homo sapiens | Homo sapiens | Moderate | NCBI |
| Life Stage | Evidence |
|---|---|
| All life stages | Moderate |
| Sex | Evidence |
|---|---|
| Unspecific | Moderate |
Prolonged ROS induces inflammation and tissue damage in Homo sapiens (Vallée & Lecarpentier, 2018).
Key Event Relationship Description
ROS production causes tissue damage (Gao, Zhou, Lin, Paus, & Yue, 2019). ROS production is involved in Wnt-driven tumorigenesis (Myant et al., 2013). The prolonged ROS induces inflammation leading to carcinogenesis (Vallée & Lecarpentier,2018). Injury causes the Porcupine-induced Wnt secretion (Saha et al., 2016).
Evidence Supporting this KER
Biological PlausibilitySustained ROS increase caused by/causes DNA damage, which will alter several signaling pathways including Wnt signaling.
Macrophages accumulate into injured tissue to recover the tissue damage, which may be followed by porcupine-induced Wnt secretion. ROS stimulate inflammatory factor production and Wnt/beta-catenin signaling (Vallée & Lecarpentier, 2018).
Empirical EvidenceProduction of ROS by DNA double-strand break causes tissue damages (Gao et al., 2019).
ROS signaling induces Wnt/beta-catenin signaling (Pérez, Taléns-Visconti, Rius-Pérez, Finamor, & Sastre, 2017).
The balance of ROS signaling is important, and dual effects of ROS should be taken in consideration. The ROS may enhance Wnt/beta-catenin proliferating pathways to promote tumorigenesis, while ROS may disrupt tumor progression by different pro-apoptotic mechanisms (Pérez et al., 2017). It is also known that Wnt signaling induces ROS signaling (Cheung et al., 2016). Wnt/beta-catenin signaling control by ROS needs to be further investigated (Caliceti, Nigro, Rizzo, & Ferrari, 2014).
Quantitative Understanding of the Linkage
Response-response relationshipROS induces inflammatory responses (Bhattacharyya, Chattopadhyay, Mitra, & Crowe, 2014). Oxidant induces ROS generation and p38 MAPK activation in macrophages (Conway & Kinter, 2006). ROS induce tissue damage in cardiac myocytes (Miller & Cheung, 2016; Yang et al., 2006).
For the colony formation assay, cells were treated with 400 microM/L H2O2 for 1 week, where the medium was changed every three days (Wang et al., 2019).
GPX2, an activator of Wnt/beta-catenin signaling, is identified as a key regulator of intracellular H2O2 levels and an inhibitor of apoptosis (Wang et al., 2019).
The reduction in ROS levels in the human serum albumin-treated cerebral ischemia/reperfusion-induced injury may be mediated by Wnt/betacatenin signaling (Tang, Shen, Zhang, Yang, & Liu, 2019).
References
Bhattacharyya, A., Chattopadhyay, R., Mitra, S., & Crowe, S. E. (2014). Oxidative stress: an essential factor in the pathogenesis of gastrointestinal mucosal diseases. Physiological reviews, 94(2), 329-354. doi:10.1152/physrev.00040.2012
Caliceti, C., Nigro, P., Rizzo, P., & Ferrari, R. (2014). ROS, Notch, and Wnt signaling pathways: crosstalk between three major regulators of cardiovascular biology. BioMed research international, 2014, 318714-318714. doi:10.1155/2014/318714
Cheung, E. C., Lee, P., Ceteci, F., Nixon, C., Blyth, K., Sansom, O. J., & Vousden, K. H. (2016). Opposing effects of TIGAR- and RAC1-derived ROS on Wnt driven proliferation in the mouse intestine. Genes & development, 30(1), 52-63. doi:10.1101/gad.271130.115
Conway, J. P., & Kinter, M. (2006). Dual role of peroxiredoxin I in macrophage-derived foam cells. The Journal of biological chemistry, 281(38), 27991-28001. doi:10.1074/jbc.M605026200
Gao, Q., Zhou, G., Lin, S.-J., Paus, R., & Yue, Z. (2019). How chemotherapy and radiotherapy damage the tissue: Comparative biology lessons from feather and hair models. Experimental dermatology, 28(4), 413-418. doi:10.1111/exd.13846
Miller, B. A., & Cheung, J. Y. (2016). TRPM2 protects against tissue damage following oxidative stress and ischaemia-reperfusion. The Journal of physiology, 594(15), 4181-4191. doi:10.1113/JP270934
Myant, K. B., Cammareri, P., McGhee, E. J., Ridgway, R. A., Huels, D. J., Cordero, J. B., . . . Sansom, O. J. (2013). ROS production and NF-κB activation triggered by RAC1 facilitate WNT-driven intestinal stem cell proliferation and colorectal cancer initiation. Cell stem cell, 12(6), 761-773. doi:10.1016/j.stem.2013.04.006
Pérez, S., Taléns-Visconti, R., Rius-Pérez, S., Finamor, I., & Sastre, J. (2017). Redox signaling in the gastrointestinal tract. Free radical biology & medicine, 104, 75-103. doi:10.1016/j.freeradbiomed.2016.12.048
Saha, S., Aranda, E., Hayakawa, Y., Bhanja, P., Atay, S., Brodin, N. P., . . . Pollard, J. W. (2016). Macrophage-derived extracellular vesicle-packaged WNTs rescue intestinal stem cells and enhance survival after radiation injury. Nature Communications, 7, 13096-13096. doi:10.1038/ncomms13096
Tang, Y., Shen, J., Zhang, F., Yang, F.-Y., & Liu, M. (2019). Human serum albumin attenuates global cerebral ischemia/reperfusion-induced brain injury in a Wnt/β- Catenin/ROS signaling-dependent manner in rats. Biomedicine & pharmacotherapy = Biomedecine & pharmacotherapie, 115, 108871-108871. doi:10.1016/j.biopha.2019.108871
Vallée, A., & Lecarpentier, Y. (2018). Crosstalk Between Peroxisome Proliferator-Activated Receptor Gamma and the Canonical WNT/β-Catenin Pathway in Chronic Inflammation and Oxidative Stress During Carcinogenesis. Frontiers in immunology, 9, 745-745. doi:10.3389/fimmu.2018.00745
Wang, Y., Cao, P., Alshwmi, M., Jiang, N., Xiao, Z., Jiang, F., . . . Li, S. (2019). GPX2 suppression of H(2)O(2) stress regulates cervical cancer metastasis and apoptosis via activation of the β-catenin-WNT pathway. OncoTargets and therapy, 12, 6639-6651. doi:10.2147/OTT.S208781
Yang, K. T., Chang, W. L., Yang, P. C., Chien, C. L., Lai, M. S., Su, M. J., & Wu, M. L. (2006). Activation of the transient receptor potential M2 channel and poly(ADP-ribose) polymerase is involved in oxidative stress-induced cardiomyocyte death. Cell Death & Differentiation, 13(10), 1815-1826. doi:10.1038/sj.cdd.4401813
Relationship: 2070: Increase, porcupine-induced Wnt secretion and Wnt signaling activation leads to beta-catenin activation
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Increase in reactive oxygen species (ROS) leading to human treatment-resistant gastric cancer | adjacent | Moderate | Moderate |
Evidence Supporting Applicability of this Relationship
| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| Homo sapiens | Homo sapiens | High | NCBI |
| Life Stage | Evidence |
|---|---|
| All life stages | High |
| Sex | Evidence |
|---|---|
| Unspecific | High |
Wnt/beta-catenin signaling, which regulates key cellular functions including proliferation, is a highly conserved pathway through evolution (Pai et al., 2017).
Key Event Relationship Description
Secreted Wnt ligand stimulates Wnt/beta-catenin signaling, in which beta-catenin is activated. Wnt ligand binds to Frizzled receptor, which leads to GSK3beta inactivation. GSK3beta inactivation leads to beta-catenin dephosphorylation, which avoids the ubiquitination of the beta-catenin and stabilizes the beta-catenin (Clevers & Nusse, 2012). The translocation of stabilized beta-catenin induces the transcription of genes involved in proliferation (Pai et al., 2017).
Evidence Supporting this KER
Biological PlausibilityCanonical Wnt pathway consists of Wnt, GSK3beta, and beta-catenin cascade (Clevers & Nusse, 2012; Hatsell, Rowlands, Hiremath, & Cowin, 2003).
GSK3beta recruitment to LRP6 leads to form un-phosphorylated beta-catenin inducing the stabilization and translocation of the beta-catenin (MacDonald, Tamai, & He, 2009).
Stabilized beta-catenin accumulates in cytosol and translocates into the nucleus leading to beta-catenin activation (MacDonald et al., 2009).
Empirical Evidence[Incidence concordance]
Dishevelled (DVL), a positive regulator of Wnt signaling, form the complex with FZD and lead to trigger the Wnt signaling together with Wnt coreceptor low-density lipoprotein (LDL) receptor-related protein 6 (LRP6) (Clevers & Nusse, 2012; Jiang, Charlat, Zamponi, Yang, & Cong, 2015). Wnt binds to FZD and activates the Wnt signaling (Clevers & Nusse, 2012; Janda, Waghray, Levin, Thomas, & Garcia, 2012; Nile, Mukund, Stanger, Wang, & Hannoush, 2017). Wnt binding towards FZD induces the formation of the protein complex with LRP5/6 and DVL, leading to the downstream signaling activation including beta-catenin (Clevers & Nusse, 2012).
Uncertainties and InconsistenciesSome Wnt ligands bind to FZD, leading to Wnt/beta-catenin signaling inactivation. DVL, a positive regulator of Wnt signaling, has a controversial role to promote Wnt receptor degradation (Jiang et al., 2015). DVL-dependent regulation of FZD level is involved in mTORC1 signaling suppression via Wnt/beta-catenin signaling (Zeng et al., 2018)
GSK3beta phosphorylates LRP6 as well as remaining GSK3 beta phosphorylates beta-catenin which would be ubiquitinated and degradated (MacDonald et al., 2009).
Quantitative Understanding of the Linkage
Response-response relationshipWnt3 promotes proliferation and survival in HUVECs (Shen et al., 2018).
GSK3beta inhibition by 1 uM of SB216763 or 5 uM of BRD3731 results in the decreased phosphorylation and stabilization of beta-catenin (Stump et al., 2019). The level of beta-catenin is increased by the inhibition of GSK3beta kinase activity (Stump et al., 2019). GSK3beta inhibition by small interference RNA (siRNA) of GSK3beta results in the decreased phosphorylation and increased expression of beta-catenin (Stump et al., 2019).
Time-scaleFZD7 enhances the activity of canonical Wnt/beta-catenin signaling with the treatment of WNT3A for 1 to 6 hrs (Cao et al., 2017). The treatment with SB216763 or BRD3731, GSK3beta inhibitors, decreases phosphorylated beta-catenin and increased beta-catenin expression in 48 hours (Stump et al., 2019). The cells are treated with GSK3beta small interference RNA (siRNA) for 48 hours to silence the expression of GSK3beta, which results in the activation of beta-catenin pathway (Stump et al., 2019).
Known modulating factorsFZD5 can activate WNT3A/beta-catenin signaling in a dose-dependent manner (Hua et al., 2018). The increase in FZD5 protein enhances cell response to WNT3A. (Hua et al., 2018). LRP5 can augment WNT3A/beta-catenin signaling in a dose-dependent manner (Hua et al., 2018). The binding of Wnt and FZD induce the formation of the protein complex with the Dvl, Axin, CK1 GSK3, beta-catenin and APC to induce the beta-catenin translocation into the nucleus (Clevers & Nusse, 2012).
Known Feedforward/Feedback loops influencing this KERBeta-catenin is required and sufficient for the sequestration of GSK3 in acidic cytoplasmic endosomes (Taelman et al., 2010). Beta-catenin, of which level increases in Wnt signaling, facilitates GSK3 sequestration leading to feed-forward loop formation (Taelman et al., 2010). The Wnt ligand is antagonized with secreted Frizzled-related proteins (sFRPs) and Wnt inhibitory protein (WIF), both of which can bind Wnts and inhibit interactions between WNT and FZD (Bovolenta, Esteve, Ruiz, Cisneros, & Lopez-Rios, 2008; Clevers & Nusse, 2012). The Dickkopf 1 (DKK1) can disrupts Wnt-induced FZD-LRP6 complex formation (Clevers & Nusse, 2012; Ellwanger et al., 2008; Semenov, Zhang, & He, 2008).
References
Bovolenta, P., Esteve, P., Ruiz, J. M., Cisneros, E., & Lopez-Rios, J. (2008). Beyond Wnt inhibition: new functions of secreted Frizzled-related proteins in development and disease. J Cell Sci, 121(Pt 6), 737-746. doi:10.1242/jcs.026096
Cao, T. T., Xiang, D., Liu, B. L., Huang, T. X., Tan, B. B., Zeng, C. M., . . . Fu, L. (2017). FZD7 is a novel prognostic marker and promotes tumor metastasis via WNT and EMT signaling pathways in esophageal squamous cell carcinoma. Oncotarget, 8(39), 65957-65968. doi:10.18632/oncotarget.19586
Clevers, H., & Nusse, R. (2012). Wnt/beta-catenin signaling and disease. Cell, 149(6), 1192-1205. doi:10.1016/j.cell.2012.05.012
Ellwanger, K., Saito, H., Clement-Lacroix, P., Maltry, N., Niedermeyer, J., Lee, W. K., . . . Niehrs, C. (2008). Targeted disruption of the Wnt regulator Kremen induces limb defects and high bone density. Mol Cell Biol, 28(15), 4875-4882. doi:10.1128/MCB.00222-08
Hatsell, S., Rowlands, T., Hiremath, M., & Cowin, P. (2003). Beta-catenin and Tcfs in mammary development and cancer. J Mammary Gland Biol Neoplasia, 8(2), 145-158. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/14635791
Hua, Y., Yang, Y., Li, Q., He, X., Zhu, W., Wang, J., & Gan, X. (2018). Oligomerization of Frizzled and LRP5/6 protein initiates intracellular signaling for the canonical WNT/beta-catenin pathway. J Biol Chem, 293(51), 19710-19724. doi:10.1074/jbc.RA118.004434
Janda, C. Y., Waghray, D., Levin, A. M., Thomas, C., & Garcia, K. C. (2012). Structural basis of Wnt recognition by Frizzled. Science, 337(6090), 59-64. doi:10.1126/science.1222879
Jiang, X., Charlat, O., Zamponi, R., Yang, Y., & Cong, F. (2015). Dishevelled promotes Wnt receptor degradation through recruitment of ZNRF3/RNF43 E3 ubiquitin ligases. Mol Cell, 58(3), 522-533. doi:10.1016/j.molcel.2015.03.015
MacDonald, B. T., Tamai, K., & He, X. (2009). Wnt/beta-catenin signaling: components, mechanisms, and diseases. Dev Cell, 17(1), 9-26. doi:10.1016/j.devcel.2009.06.016
Nile, A. H., Mukund, S., Stanger, K., Wang, W., & Hannoush, R. N. (2017). Unsaturated fatty acyl recognition by Frizzled receptors mediates dimerization upon Wnt ligand binding. Proc Natl Acad Sci U S A, 114(16), 4147-4152. doi:10.1073/pnas.1618293114
Pai SG, Carneiro BA, Mota JM, Costa R, Leite CA, Barroso-Sousa R, Kaplan JB, Chae YK, Giles FJ. Wnt/beta-catenin pathway: modulating anticancer immune response. J Hematol Oncol. 2017 May 5;10(1):101. doi: 10.1186/s13045-017-0471-6. PMID: 28476164; PMCID: PMC5420131.
Semenov, M. V., Zhang, X., & He, X. (2008). DKK1 antagonizes Wnt signaling without promotion of LRP6 internalization and degradation. J Biol Chem, 283(31), 21427-21432. doi:10.1074/jbc.M800014200
Shen, M., Bai, D., Liu, B., Lu, X., Hou, R., Zeng, C., . . . Yin, T. (2018). Dysregulated Txnip-ROS-Wnt axis contributes to the impaired ischemic heart repair in diabetic mice. Biochimica et biophysica acta. Molecular basis of disease, 1864(12), 3735-3745. doi:10.1016/j.bbadis.2018.09.029
Stump, B., Shrestha, S., Lamattina, A. M., Louis, P. H., Cho, W., Perrella, M. A., . . . El-Chemaly, S. (2019). Glycogen synthase kinase 3-beta inhibition induces lymphangiogenesis through beta-catenin-dependent and mTOR-independent pathways. PLoS One, 14(4), e0213831. doi:10.1371/journal.pone.0213831
Taelman, V. F., Dobrowolski, R., Plouhinec, J. L., Fuentealba, L. C., Vorwald, P. P., Gumper, I., . . . De Robertis, E. M. (2010). Wnt signaling requires sequestration of glycogen synthase kinase 3 inside multivesicular endosomes. Cell, 143(7), 1136-1148. doi:10.1016/j.cell.2010.11.034
Zeng, H., Lu, B., Zamponi, R., Yang, Z., Wetzel, K., Loureiro, J., . . . Cong, F. (2018). mTORC1 signaling suppresses Wnt/beta-catenin signaling through DVL-dependent regulation of Wnt receptor FZD level. Proc Natl Acad Sci U S A, 115(44), E10362-E10369. doi:10.1073/pnas.1808575115
Relationship: 3199: beta-catenin activation leads to EMT
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Increase in reactive oxygen species (ROS) leading to human treatment-resistant gastric cancer | adjacent | Moderate | Moderate |
Evidence Supporting Applicability of this Relationship
| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| Homo sapiens | Homo sapiens | High | NCBI |
| Life Stage | Evidence |
|---|---|
| All life stages | High |
| Sex | Evidence |
|---|---|
| Unspecific | High |
- The inhibition of c-MET decreases the expression of beta-catenin and Snail in human diffuse-type gastric cancer (Homo sapiens) (Sohn et al., 2019).
- The treatment with garcinol decreases the expression of beta-catenin and ZEB1/ZEB2 in human breast cancer cells (Homo sapiens) (Ahmad et al., 2012).
- Zeb1 activation leads to EMT via Prex1 activation in NCH421k, NCH441, and NCH644 human glioblastoma model cells (Homo sapiens) (Rosmaninho et al., 2018).
- Zeb1 siRNA induced the suppression of EMT in SGC-7901 human gastric cancer cell line (Homo sapiens) (Xue et al., 2019). Snail induces EMT in SAS and HSC-4 human head and neck squamous cancer cells (Homo sapiens) (Ota et al., 2016).
- Snail induces EMT in B16-F10 murine melanoma cells (Mus musculus) (Kudo-Saito, Shirako, Takeuchi, & Kawakami, 2009; Wang, Shi, Chai, Ying, & Zhou, 2013).
- Twist1 is related to EMT in MCF-7 and MDA-MB-231 human breast cancer cell lines (Homo sapiens) (Menendez-Menendez et al., 2019).
- Twist induces EMT in Huh7 human hepatocellular carcinoma cell lines (Homo sapiens) (Hu et al., 2019).
Key Event Relationship Description
Beta-catenin activation, of which mechanism include the stabilization of the dephosphorylated beta-catenin and translocation of beta-catenin into the nucleus, induce the formation of beta-catenin-TCF complex and transcription of transcription factors such as Snail, Zeb and Twist (Clevers & Nusse, 2012) (Ahmad et al., 2012; Pearlman, Montes de Oca, Pal, & Afaq, 2017; Sohn et al., 2019; Yang et al., 2019).
EMT-related transcription factors including Snail, ZEB and Twist are up-regulated in cancer cells (Diaz, Vinas-Castells, & Garcia de Herreros, 2014). The transcription factors such as Snail, ZEB and Twist bind to E-cadherin (CDH1) promoter and inhibit the CDH1 transcription via the consensus E-boxes (5’-CACCTG-3’ or 5’-CAGGTG-3’), which leads to EMT (Diaz et al., 2014).
Evidence Supporting this KER
Biological PlausibilityThe treatment of human gastric cancer cells with INC280, which inhibits c-MET overexpressed in diffuse-type gastric cancer with poor prognosis, shows downregulation in beta-catenin and Snail expression,(Sohn et al., 2019).
The treatment with garcinol, a polyisoprenylated benzophenone derivative that is obtained from Garcinia indica extract, induced ZEB1 and ZEB2 down-regulation, increase in phosphorylated beta-catenin, and decrease in nuclear beta-catenin in human breast cancer cells (Ahmad et al., 2012).
Sortilin, a member of the Vps10p sorting receptor family which is highly expressed in high-grade malignant glioma, positively regulates GSK-3beta/beta-catenin/Twist signaling pathway in glioblastoma (Yang et al., 2019).
TM4SF1 promotes EMT via Wnt/beta-catenin/SOX2 pathway in colorectal cancer (Yang et al., 2020).
The transcription factors such as Snail, Zeb, and Twist inhibit the CDH1 expression through their binding towards the promoter of CDH1, which leads to inhibition of cell adhesion and EMT (Diaz et al., 2014)
Empirical Evidence[Dose concordance]
The inhibition of sortilin by AF38469 (a sortilin inhibitor) or small interference RNA (siRNA) results in a decrease in beta-catenin and Twist expression in human glioblastoma cells (Yang et al., 2019).
[Time concordance]
The complex of beta-catenin and TCF4 induces epithelial-mesenchymal transition (EMT)-activator ZEB (Sanchez-Tillo E et al., 2011).
[Incidence concordance]
The inhibition of c-MET, which is overexpressed in diffuse-type gastric cancer, induced an increase in phosphorylated beta-catenin, decrease in beta-catenin and Snail (Sohn et al., 2019).
The garcinol, which has an anti-cancer effect, increases phosphorylated beta-catenin, decreases beta-catenin and ZEB1/ZEB2, and inhibits EMT (Ahmad et al., 2012).
Histone deacetylase inhibitors affect EMT-related transcription factors including ZEB, Twist, and Snail (Wawruszak et al., 2019).
Snail and Zeb induces EMT and suppress E-cadherin (CDH1) (Batlle et al., 2000; Diaz et al., 2014; Peinado, Olmeda, & Cano, 2007).
Uncertainties and InconsistenciesIt is possible that the inhibition of ZEB1 and ZEB2 by garcinol treatment is caused by down-regulation of NFkappaB and Wnt/beta-catenin signaling (Ahmad et al., 2012).
The EMT is induced different transcription factors other than Zeb, Twist, and Snail, which includes E47 and KLF8 (Diaz et al., 2014).
Zeb, Twist, and Snail may activate or inactivate different genes or molecules to induce phenomena related to EMT and other phenomena other than EMT (Li & Balazsi, 2018).
Quantitative Understanding of the Linkage
Response-response relationshipThe treatment with AF38469, a sortilin inhibitor, in 0, 100, 200, 400, 800, and 1600 nM concentration inhibited beta-catenin and Twist (EMT regulator) expression dose-dependently in human glioblastoma cells (Yang et al., 2019).
Snail (SNAI1, a key transcription factor of EMT induced by beta-catenin) mRNA is methylated, and N6-methyladenosine (m6A) in its coding region (CDS) and 3’ untranslated region (3’UTR) are significantly enriched during EMT progression (Lin et al., 2019). The m6A enrichment fold of SNAI1 mRNA in EMT cells is about 2.3-fold greater than in control cells (Lin et al., 2019).
Time-scaleNuclear accumulation of beta-catenin induces endogenous ZEB1 in 15 and 30 min (Sanchez-Tillo E et al., 2011).
The treatment with 25 uM of garcinol for 48 hours induced an increase in phosphorylated beta-catenin and decreased nuclear beta-catenin protein and ZEB1/ZEB2 mRNA in human breast cancer cells (Ahmad et al., 2012).
The treatment with AF38469, a sortilin inhibitor, for 0, 2, 4, 8, 16, or 24 hours shows that the expression of beta-catenin and Twist decrease in 8 hours followed by the subsequent decrease in 16 and 24 hours in human glioblastoma cells (Yang et al., 2019).
Snail (SNAI1) transfection for 48 hours induces the repression of E-cadherin (CDH1) protein expression (Lin et al., 2019).
SNAI1 mRNA in polysome is up-regulated in EMT-undergoing HeLa cells treated with 10 ng/ml of TGF-beta for 3 days compared with control cells (Lin et al., 2019).
Known modulating factorsThe proto-oncogene MET regulates beta-catenin and Snail expression (Sohn et al., 2019).
The inhibition of GSK3beta by SB216763 induced expression of beta-catenin and Twist, as well as mesenchymal markers such as N-cadherin, vimentin, and MMP9 (Yang et al., 2019).
The decrease in E-cadherin (CDH1), a cell adhesion molecule, is related to EMT (Diaz et al., 2014).
Methyltransferase-like 3 (METTL3) modulates methylation of Snail (SNAI1) mRNA and EMT (Lin et al., 2019).
The binding of beta-catenin to members of the TCF/LEF family transcription factors increase gene expression related to EMT such as Twist and decrease E-cadherin protein expression (Qualtrough, Rees, Speight, Williams, & Paraskeva, 2015).
Known Feedforward/Feedback loops influencing this KER
The inhibited expression of phosphorylated GSK3beta, beta-catenin, and Twist by sortilin inhibition is reversed by GSK3beta inhibition. Furthermore, twist overexpression by lentivirus increased the inhibited expression of N-cadherin, MMP9, and vimentin and reverses the inhibitory effect of AF38469 on sortilin, which suggests that sortilin induces glioblastoma invasion mainly via GSK3beta/beta-catenin/Twist induced mesenchymal transition (Yang et al., 2019).
The inhibition of Hedgehog signaling pathway with cyclopamine reduces beta-catenin-TCF transcriptional activity, decreases the Twist expression, induces E-cadherin expression, and inhibits EMT (Qualtrough et al., 2015).
References
Ahmad, A., Sarkar, S. H., Bitar, B., Ali, S., Aboukameel, A., Sethi, S., . . . Sarkar, F. H. (2012). Garcinol regulates EMT and Wnt signaling pathways in vitro and in vivo, leading to anticancer activity against breast cancer cells. Mol Cancer Ther, 11(10), 2193-2201. doi:10.1158/1535-7163.MCT-12-0232-T
Batlle, E., Sancho, E., Francí, C., Domínguez, D., Monfar, M., Baulida, J., & García de Herreros, A. (2000). The transcription factor Snail is a repressor of E-cadherin gene expression in epithelial tumour cells. Nature Cell Biology, 2(2), 84-89. doi:10.1038/35000034
Clevers, H., & Nusse, R. (2012). Wnt/beta-catenin signaling and disease. Cell, 149(6), 1192-1205. doi:10.1016/j.cell.2012.05.012
Diaz, V. M., Vinas-Castells, R., & Garcia de Herreros, A. (2014). Regulation of the protein stability of EMT transcription factors. Cell Adh Migr, 8(4), 418-428. doi:10.4161/19336918.2014.969998
Hu, B., Cheng, J. W., Hu, J. W., Li, H., Ma, X. L., Tang, W. G., . . . Yang, X. R. (2019). KPNA3 Confers Sorafenib Resistance to Advanced Hepatocellular Carcinoma via TWIST Regulated Epithelial-Mesenchymal Transition. Journal of Cancer, 10(17), 3914-3925. doi:10.7150/jca.31448
Kudo-Saito, C., Shirako, H., Takeuchi, T., & Kawakami, Y. (2009). Cancer Metastasis Is Accelerated through Immunosuppression during Snail-Induced EMT of Cancer Cells. Cancer Cell, 15(3), 195-206. doi:10.1016/j.ccr.2009.01.023
Li, C., & Balazsi, G. (2018). A landscape view on the interplay between EMT and cancer metastasis. NPJ Syst Biol Appl, 4, 34. doi:10.1038/s41540-018-0068-x
Lin, X., Chai, G., Wu, Y., Li, J., Chen, F., Liu, J., . . . Wang, H. (2019). RNA m(6)A methylation regulates the epithelial mesenchymal transition of cancer cells and translation of Snail. Nat Commun, 10(1), 2065. doi:10.1038/s41467-019-09865-9
Menendez-Menendez, J., Hermida-Prado, F., Granda-Diaz, R., Gonzalez, A., Garcia-Pedrero, J. M., Del-Rio-Ibisate, N., . . . Martinez-Campa, C. (2019). Deciphering the Molecular Basis of Melatonin Protective Effects on Breast Cells Treated with Doxorubicin: TWIST1 a Transcription Factor Involved in EMT and Metastasis, a Novel Target of Melatonin. Cancers (Basel), 11(7). doi:10.3390/cancers11071011
Ota, I., Masui, T., Kurihara, M., Yook, J. I., Mikami, S., Kimura, T., . . . Kitahara, T. (2016). Snail-induced EMT promotes cancer stem cell-like properties in head and neck cancer cells. Oncol Rep, 35(1), 261-266. doi:10.3892/or.2015.4348
Pearlman, R. L., Montes de Oca, M. K., Pal, H. C., & Afaq, F. (2017). Potential therapeutic targets of epithelial-mesenchymal transition in melanoma. Cancer Lett, 391, 125-140. doi:10.1016/j.canlet.2017.01.029
Peinado, H., Olmeda, D., & Cano, A. (2007). Snail, Zeb and bHLH factors in tumour progression: an alliance against the epithelial phenotype? Nat Rev Cancer, 7(6), 415-428. doi:10.1038/nrc2131
Qualtrough, D., Rees, P., Speight, B., Williams, A. C., & Paraskeva, C. (2015). The Hedgehog Inhibitor Cyclopamine Reduces beta-Catenin-Tcf Transcriptional Activity, Induces E-Cadherin Expression, and Reduces Invasion in Colorectal Cancer Cells. Cancers (Basel), 7(3), 1885-1899. doi:10.3390/cancers7030867
Rosmaninho, P., Mükusch, S., Piscopo, V., Teixeira, V., Raposo, A. A., Warta, R., . . . Castro, D. S. (2018). Zeb1 potentiates genome-wide gene transcription with Lef1 to promote glioblastoma cell invasion. The EMBO Journal, 37(15), e97115. doi:10.15252/embj.201797115
Sanchez-Tillo E, de Barrios O, Siles L, Cuatrecasas M, Castells A, Postigo A. beta-catenin/TCF4 complex induces the epithelial-to-mesenchymal transition (EMT)-activator ZEB1 to regulate tumor invasiveness. Proc Natl Acad Sci U S A, 2011;108(48):19204-9.
Sohn, S. H., Kim, B., Sul, H. J., Kim, Y. J., Kim, H. S., Kim, H., . . . Zang, D. Y. (2019). INC280 inhibits Wnt/beta-catenin and EMT signaling pathways and its induce apoptosis in diffuse gastric cancer positive for c-MET amplification. BMC Res Notes, 12(1), 125. doi:10.1186/s13104-019-4163-x
Tang Q, Chen J, Di Z, Yuan W, Zhou Z, Liu Z, Han S, Liu Y, Ying G, Shu X, Di M. TM4SF1 promotes EMT and cancer stemness via the Wnt/β-catenin/SOX2 pathway in colorectal cancer. J Exp Clin Cancer Res. 2020 Nov 5;39(1):232. doi: 10.1186/s13046-020-01690-z. PMID: 33153498; PMCID: PMC7643364.
Wang, Y., Shi, J., Chai, K., Ying, X., & Zhou, B. P. (2013). The Role of Snail in EMT and Tumorigenesis. Current cancer drug targets, 13(9), 963-972. doi: 10.2174/15680096113136660102
Wawruszak, A., Kalafut, J., Okon, E., Czapinski, J., Halasa, M., Przybyszewska, A., . . . Stepulak, A. (2019). Histone Deacetylase Inhibitors and Phenotypical Transformation of Cancer Cells. Cancers (Basel), 11(2). doi:10.3390/cancers11020148
Xue, Y., Zhang, L., Zhu, Y., Ke, X., Wang, Q., & Min, H. (2019). Regulation of Proliferation and Epithelial-to-Mesenchymal Transition (EMT) of Gastric Cancer by ZEB1 via Modulating Wnt5a and Related Mechanisms. Medical science monitor : international medical journal of experimental and clinical research, 25, 1663-1670. doi:10.12659/MSM.912338
Yang, W., Wu, P. F., Ma, J. X., Liao, M. J., Wang, X. H., Xu, L. S., . . . Yi, L. (2019). Sortilin promotes glioblastoma invasion and mesenchymal transition through GSK-3beta/beta-catenin/twist pathway. Cell Death Dis, 10(3), 208. doi:10.1038/s41419-019-1449-9
Relationship: 3200: EMT leads to Resistant gastric cancer
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Increase in reactive oxygen species (ROS) leading to human treatment-resistant gastric cancer | adjacent | Moderate | Moderate |
Evidence Supporting Applicability of this Relationship
| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| Homo sapiens | Homo sapiens | High | NCBI |
| Life Stage | Evidence |
|---|---|
| All life stages | High |
| Sex | Evidence |
|---|---|
| Unspecific | High |
EMT induces cancer invasion, metastasis (Homo sapiens)(P. Zhang et al., 2015).
EMT is related to cancer drug resistance in MCF-7 human breast cancer cells (Homo sapiens)(B. Du & Shim, 2016).
Key Event Relationship Description
Some population of the cells exhibiting EMT demonstrates the feature of cancer stem cells (CSCs), which are related to cancer malignancy (Shibue & Weinberg, 2017; Shihori Tanabe, 2015a, 2015b; Tanabe, Aoyagi, Yokozaki, & Sasaki, 2015).
EMT phenomenon is related to cancer metastasis and cancer therapy resistance (Smith & Bhowmick, 2016; Tanabe, 2013). The increased expression of enzymes that degrade the extracellular matrix components and the decrease in adhesion to the basement membrane in EMT induces the cell to escape from the basement membrane and metastasis (Smith & Bhowmick, 2016). Morphological changes observed during EMT are associated with therapy resistance (Smith & Bhowmick, 2016).
Evidence Supporting this KER
Biological PlausibilityThe morphological and physiological changes associated with EMT are involved in invasiveness and drug resistance (Shibue & Weinberg, 2017). The EMT-activated particular carcinoma cells in primary tumors invade the surrounding stroma (Shibue & Weinberg, 2017). The EMT –activated carcinoma cells interact with the surrounding extracellular matrix protein to induce focal adhesion kinase and extracellular signal-related kinase activation, followed by the transforming growth factor-beta (TGFbeta) and canonical and/or noncanonical Wnt pathways to induce cancer stem cell (CSC) properties which contribute to the drug resistance (Shibue & Weinberg, 2017).
EMT-associated down-regulation of multiple apoptotic signaling pathways induces drug efflux and slows cell proliferation to induce the general resistance of carcinoma cells to anti-cancer drugs (Shibue & Weinberg, 2017).
Snail, an EMT-related transcription factor, induces the expression of the AXL receptor tyrosine kinase, which enables the cancer cells to survive by the activation of AXL signaling triggered by the binding of its ligand growth arrest-specific protein 6 (GAS6)(Shibue & Weinberg, 2017).
The EMT-activated cells evade the lethal effect of cytotoxic T cells, which include the elevated expression of programmed cell death 1 ligand (PD-L1) which binds to the programmed cell death protein 1 (PD-1) inhibitory immune-checkpoint receptor on the cell surface of cytotoxic T cells (Shibue & Weinberg, 2017).
Empirical EvidenceIncidence concordance
Slug/Snai2, a ces-1-related zinc finger transcription factor gene, confers resistance to p53-mediated apoptosis of hematopoietic progenitors by repressing PUMA (also known as BBC3, encoding Bcl-2-binding component 3) (Inukai et al., 1999; Shibue & Weinberg, 2017; W.-S. Wu et al., 2005).
EMT activation induces the expression of multiple members of the ATP-binding cassette (ABC) transporter family, which results in the resistance to doxorubicin (Saxena, Stephens, Pathak, & Rangarajan, 2011; Shibue & Weinberg, 2017)
TGFbeta-1 induced EMT results in the acquisition of cancer stem cell (CSC) like properties (Pirozzi et al., 2011; Shibue & Weinberg, 2017).
Snail-induced EMT induces cancer metastasis and resistance to dendritic cell-mediated immunotherapy (Kudo-Saito, Shirako, Takeuchi, & Kawakami, 2009).
Zinc finger E-box-binding homeobox (ZEB1)-induced EMT results in the relief of miR-200-mediated repression of programmed cell death 1 ligand (PD-L1) expression, a major inhibitory ligand for the programmed cell death protein (PD-1) immune-checkpoint protein on CD8+ cytotoxic T lymphocyte (CTL), subsequently the CD8+ T cell immunosuppression and metastasis (Chen et al., 2014).
Uncertainties and InconsistenciesThe reversing process of EMT, which names as a mesenchymal-epithelial transition (MET), maybe one of the candidates for the anti-cancer therapy, where the plasticity of the cell phenotype is of importance and under investigation (Shibue & Weinberg, 2017).
Quantitative Understanding of the Linkage
Response-response relationshipInduction of EMT by TGFbeta and Twist increases the gene expression of EMT markers such as Snail, Vimentin, N-cadherin, and ABC transporters including ABCA3, ABCC1, ABCC3, and ABCC10 (Saxena et al., 2011).
Human mammary epithelial cells (HMLE) stably expressing Twist, FOXC2 or Snail demonstrates the increased cell viability compared to control HMLE in the treatment with about 0.3, 3, 30 mM of doxorubicin, dose-dependently (Saxena et al., 2011).
Time-scaleThe treatment with doxorubicin for 48 hours demonstrates the increase in the cell viability in Twist/FOXC2/Snail overexpressed HMLE compared to control HMLE (Saxena et al., 2011).
The inhibition of Twist or Zeb1 with small interference RNA (siRNA) induced the inhibition of cell viability compared to control MDAMB231 cells treated with doxorubicin for 48 hours (Saxena et al., 2011).
Known modulating factorsABC transporters that are related to drug resistance are overexpressed in the EMT-activated cells (Saxena et al., 2011). The expression of PD-L1, which binds to the PD-1 on the cytotoxic T cells, is up-regulated in EMT-activated cells, which results in the inhibition of cancer immunity and the resistance to cancer therapy (Shibue & Weinberg, 2017).
Known Feedforward/Feedback loops influencing this KERThe investigation of EMT-CSC relations is important to understand the relationship between EMT and cancer malignancy. Non-CSCs in cancer can spontaneously undergo EMT and dedifferentiate into new CSC, subsequently induce the regeneration of tumorigenic potential (Marjanovic, Weinberg, & Chaffer, 2013; Shibue & Weinberg, 2017).
The plastic CSC theory demonstrates the bidirectional conversions between non-CSCs and CSCs, which may contribute to the acquisition of cancer malignancy in EMT-activated cells (Marjanovic et al., 2013).
References
Chen, L., Gibbons, D. L., Goswami, S., Cortez, M. A., Ahn, Y.-H., Byers, L. A., . . . Qin, F. X.-F. (2014). Metastasis is regulated via microRNA-200/ZEB1 axis control of tumour cell PD-L1 expression and intratumoral immunosuppression. Nature communications, 5, 5241-5241. doi:10.1038/ncomms6241
Du, B., & Shim, J. S. (2016). Targeting Epithelial-Mesenchymal Transition (EMT) to Overcome Drug Resistance in Cancer. Molecules, 21(7). doi:10.3390/molecules21070965
Inukai, T., Inoue, A., Kurosawa, H., Goi, K., Shinjyo, T., Ozawa, K., . . . Look, A. T. (1999). SLUG, a ces-1-Related Zinc Finger Transcription Factor Gene with Antiapoptotic Activity, Is a Downstream Target of the E2A-HLF Oncoprotein. Molecular Cell, 4(3), 343-352. doi:https://doi.org/10.1016/S1097-2765(00)80336-6
Kudo-Saito, C., Shirako, H., Takeuchi, T., & Kawakami, Y. (2009). Cancer Metastasis Is Accelerated through Immunosuppression during Snail-Induced EMT of Cancer Cells. Cancer Cell, 15(3), 195-206. doi:https://doi.org/10.1016/j.ccr.2009.01.023
Marjanovic, N. D., Weinberg, R. A., & Chaffer, C. L. (2013). Cell plasticity and heterogeneity in cancer. Clinical chemistry, 59(1), 168-179. doi:10.1373/clinchem.2012.184655
Pirozzi, G., Tirino, V., Camerlingo, R., Franco, R., La Rocca, A., Liguori, E., . . . Rocco, G. (2011). Epithelial to mesenchymal transition by TGFβ-1 induction increases stemness characteristics in primary non small cell lung cancer cell line. PLoS One, 6(6), e21548-e21548. doi:10.1371/journal.pone.0021548
Saxena, M., Stephens, M. A., Pathak, H., & Rangarajan, A. (2011). Transcription factors that mediate epithelial-mesenchymal transition lead to multidrug resistance by upregulating ABC transporters. Cell death & disease, 2(7), e179-e179. doi:10.1038/cddis.2011.61
Shibue, T., & Weinberg, R. A. (2017). EMT, CSCs, and drug resistance: the mechanistic link and clinical implications. Nat Rev Clin Oncol, 14(10), 611-629. doi:10.1038/nrclinonc.2017.44
Smith, B. N., & Bhowmick, N. A. (2016). Role of EMT in Metastasis and Therapy Resistance. J Clin Med, 5(2). doi:10.3390/jcm5020017
Tanabe, S. (2013). Perspectives of gene combinations in phenotype presentation. World journal of stem cells, 5(3), 61-67. doi:10.4252/wjsc.v5.i3.61
Tanabe, S. (2015a). Origin of cells and network information. World journal of stem cells, 7(3), 535-540. doi:10.4252/wjsc.v7.i3.535
Tanabe, S. (2015b). Signaling involved in stem cell reprogramming and differentiation. World journal of stem cells, 7(7), 992-998. doi:10.4252/wjsc.v7.i7.992
Tanabe, S., Aoyagi, K., Yokozaki, H., & Sasaki, H. (2015). Regulated genes in mesenchymal stem cells and gastric cancer. World journal of stem cells, 7(1), 208-222. doi:10.4252/wjsc.v7.i1.208
Wu, W.-S., Heinrichs, S., Xu, D., Garrison, S. P., Zambetti, G. P., Adams, J. M., & Look, A. T. (2005). Slug Antagonizes p53-Mediated Apoptosis of Hematopoietic Progenitors by Repressing puma. Cell, 123(4), 641-653. doi:https://doi.org/10.1016/j.cell.2005.09.029
Zhang, P., Sun, Y., & Ma, L. (2015). ZEB1: at the crossroads of epithelial-mesenchymal transition, metastasis and therapy resistance. Cell Cycle, 14(4), 481-487. doi:10.1080/15384101.2015.1006048