AOP ID and Title:
Graphical Representation

Status
| Author status | OECD status | OECD project | SAAOP status |
|---|---|---|---|
| Under development: Not open for comment. Do not cite |
Summary of the AOP
Events
Molecular Initiating Events (MIE), Key Events (KE), Adverse Outcomes (AO)
| Sequence | Type | Event ID | Title | Short name |
|---|---|---|---|---|
| MIE | 279 | Thyroperoxidase, Inhibition | Thyroperoxidase, Inhibition | |
| KE | 277 | Thyroid hormone synthesis, Decreased | TH synthesis, Decreased | |
| KE | 281 | Thyroxine (T4) in serum, Decreased | T4 in serum, Decreased | |
| KE | 1003 | Decreased, Triiodothyronine (T3) in serum | Decreased, Triiodothyronine (T3) in serum | |
| KE | 1877 | Altered, retinal layer structure | Altered, retinal layer structure | |
| AO | 1643 | Altered, Visual function | Altered, Visual function | |
| AO | 351 | Increased Mortality | Increased Mortality | |
| AO | 360 | Decrease, Population trajectory | Decrease, Population trajectory |
Key Event Relationships
| Upstream Event | Relationship Type | Downstream Event | Evidence | Quantitative Understanding |
|---|---|---|---|---|
| Thyroperoxidase, Inhibition | adjacent | Thyroid hormone synthesis, Decreased | High | Low |
| Thyroid hormone synthesis, Decreased | adjacent | Thyroxine (T4) in serum, Decreased | Moderate | Low |
| Thyroxine (T4) in serum, Decreased | adjacent | Decreased, Triiodothyronine (T3) in serum | Moderate | Moderate |
| Decreased, Triiodothyronine (T3) in serum | adjacent | Altered, retinal layer structure | ||
| Altered, retinal layer structure | adjacent | Altered, Visual function | ||
| Altered, Visual function | adjacent | Increased Mortality | ||
| Increased Mortality | adjacent | Decrease, Population trajectory | High | Moderate |
| Thyroperoxidase, Inhibition | non-adjacent | Thyroxine (T4) in serum, Decreased | High | Low |
Stressors
| Name | Evidence |
|---|---|
| Propylthiouracil | High |
Overall Assessment of the AOP
Domain of Applicability
Life Stage Applicability| Life Stage | Evidence |
|---|---|
| Embryo | Moderate |
| Larvae | Moderate |
| Foetal | Moderate |
| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| zebrafish | Danio rerio | NCBI |
| Sex | Evidence |
|---|---|
| Unspecific | Moderate |
Life stage applicability: This AOP currently considers the impact of reduced TH synthesis on the development of the retina which starts during embryonic development across vertebrates and continues during later life stages, e.g., until after birth in mammals and into the larval stage in fish. The focus is mainly on embryolarval/embryofoetal development. At this point, this AOP does not consider potential effects of thyroid hormone system disruption on the already developed retina during later life stages.
In order to more specifically evaluate the life stage applicability of the impact on thyroperoxidase inhibition on retinal layer structure and visual function leading to increased mortality, the timing of the ontogeny of the target organ needs to be matched to the timing of the ontogeny of the HPT-axis. Fish, amphibians and birds develop externally and rely on maternally transferred THs and TH machinery during the earliest stages of embryonic development.
In zebrafish, effects on retinal layer structure are typically observed at 96 or 120 hpf. By 60 hpf, the different layers of the retina can be distinguished (Morris and Fadool 2005; Schmitt and Dowling 1999) but differentiation and maturation continues until well beyond 84 hpf (Raymond and others 1995). The first thyroid follicle appears around 55 hpf and endogenous T4 production has been observed at 72 hpf (Walter and others 2019). Since thyroperoxidase is principally located in the thyroid follicles and responsible for the synthesis of TH which are released to circulation, important impacts on thyroidal TH synthesis due to thyroperoxidase inhibition are not expected before 72 hpf. This hypothesis is in line with the observation that inflation of the posterior chamber of the swim bladder appears to be unaffected by thyroperoxidase inhibition in zebrafish and fathead minnow (Nelson and others 2016; Stinckens and others 2016). Thyroperoxidase expression has however additionally been observed locally in the eyes of mice and zebrafish (Li and others 2012), suggesting a potential role of local TH synthesis in eye development before the thyroid follicles become active.
In summary, in zebrafish the formation of the retinal layers occurs before the activation of thyroidal TH synthesis but further differentiation and maturation continues until well after the onset of thyroidal TH synthesis. There is ample evidence of the impact of thyroperoxidase inhibition on retinal layer structure at the age of 5 dpf, and there is some evidence showing early effects at 48, 66 and 72 hpf (Komoike and others 2013; Reider and Connaughton 2014) suggesting the importance of local TH synthesis in the eyes. Additional mechanisms (e.g., deiodinase inhibition) could also play a role. Currently there is insufficient evidence to clearly evaluate the importance of inhibition of local TPO in the eyes versus thyroidal TPO for the development of proper retinal layer structure.
Mammals on the other hand continuously receive maternal THs via the placenta. Therefore, exposure to inhibitors of TH synthesis is expected to have an effect on the earliest phases of embryonic development by inhibiting maternal TH synthesis (REF).
Taken together, there is strong support for applicability of the current AOP to embryolarval/embryofoetal stages of vertebrates.
Taxonomic applicability: The weight of evidence supporting the first linkage of this AOP between the MIE, TPO inhibition, and the KE of decreased TH synthesis, is strong and supported by more than three decades of research in animals including humans. Several papers have measured alterations in TPO and subsequent effects on TH synthesis (Cooper et al., 1982; Cooper et al.,1983; Divi and Doerge, 1994).
Also for the next KER, it is widely accepted that TPO inhibition leads to declines in serum T4 levels in adult mammals. Strong qualitative and quantitative relationships exist between reduced TH synthesis and reduced serum T4 (Ekerot et al., 2013; Degon et al., 2008; Cooper et al., 1982; 1983; Leonard et al., 2016; Zoeller and Tan, 2007). Nevertheless, a majority of the empirical evidence comes from work with laboratory rodents, there is a large amount of supporting data from humans (with anti-hyperthyroidism drugs including propylthiouracil and methimazole), some amphibian species (e.g., frog), fish species (e.g., zebrafish and fathead minnow), and some avian species (e.g, chicken) (Cooper et al. (1982; 1983); Hornung et al. (2010); Van Herck et al. (2013); Paul et al. (2013); Nelson et al. (2016); Alexander et al. (2017); Stinckens et al. (2020)).
Although the following KER (T4 in serum decreased leads to Triiodothyronine (T3) in serum decreased) is plausibly applicable across vertebrates, too, variation can be expecteddue to feedback/compensatory mechanisms that can also differ across species. In zebrafish and fathead minnow, several studies reported the evidence for a relationship between circulating T4 and T3 levels (Nelson et al., 2016; Stinckens et al., 2020, Wang et al., 2020).
The linkage between the MIE, decreased T3 in serum, and the KE of altered retinal layer structure, is evidental in the different vertebrate classes. There is ample evidence that THs have an influence on the development of the retinal layer structure. Although there are some differences in eye structure between species, it is known that the retina follows the typical organisation of vertebrates. Within vertebrates, it consists of several layers of RPE, photoreceptors, neurons and choroid. It is plausible to assume that TH levels are important for healthy eye development in all vertebrates.
Thyroid hormone receptors have a general function in different cell types of the vertebrate retina, they mediate specific events in retinal and photoreceptor development. The decrease of TH levels can lead to disturbances of the retinal layers, as shown by studies in various vertebrates such as fish species, rats, mice and humans (Baumann (2016), Komoike et al. (2013), Besson et al. (2020), Gamborino (2000), Houbrechts (2016), (Li et al. 2021)).
References
Komoike Y, Matsuoka M, Kosaki K. 2013. Potential Teratogenicity of Methimazole: Exposure of Zebrafish Embryos to Methimazole Causes Similar Developmental Anomalies to Human Methimazole Embryopathy. Birth Defects Research Part B-Developmental and Reproductive Toxicology 98(3):222-229.
Li Z, Ptak D, Zhang L, Walls EK, Zhong W, Leung YF. 2012. Phenylthiourea Specifically Reduces Zebrafish Eye Size. Plos One 7(6).
Morris AC, Fadool JM. 2005. Studying rod photoreceptor development in zebrafish. Physiology & Behavior 86(3):306-313.
Nelson K, Schroeder A, Ankley G, Blackwell B, Blanksma C, Degitz S, Flynn K, Jensen K, Johnson R, Kahl M et al. . 2016. Impaired anterior swim bladder inflation following exposure to the thyroid peroxidase inhibitor 2-mercaptobenzothiazole part I: Fathead minnow. Aquatic Toxicology 173:192-203.
Raymond PA, Barthel LK, Curran GA. 1995. DEVELOPMENTAL PATTERNING OF ROD AND CONE PHOTORECEPTORS IN EMBRYONIC ZEBRAFISH. Journal of Comparative Neurology 359(4):537-550.
Reider M, Connaughton VP. 2014. Effects of Low-Dose Embryonic Thyroid Disruption and Rearing Temperature on the Development of the Eye and Retina in Zebrafish. Birth Defects Research Part B-Developmental and Reproductive Toxicology 101(5):347-354.
Schmitt EA, Dowling JE. 1999. Early retinal development in the zebrafish, Danio rerio: Light and electron microscopic analyses. Journal of Comparative Neurology 404(4):515-536.
Stinckens E, Vergauwen L, Schroeder A, Maho W, Blackwell B, Witters H, Blust R, Ankley G, Covaci A, Villeneuve D et al. . 2016. Impaired anterior swim bladder inflation following exposure to the thyroid peroxidase inhibitor 2-mercaptobenzothiazole part II: Zebrafish. Aquatic Toxicology 173:204-217.
Walter KM, Miller GW, Chen XP, Yaghoobi B, Puschner B, Lein PJ. 2019. Effects of thyroid hormone disruption on the ontogenetic expression of thyroid hormone signaling genes in developing zebrafish (Danio rerio). General and Comparative Endocrinology 272:20-32.
Appendix 1
List of MIEs in this AOP
Event: 279: Thyroperoxidase, Inhibition
Short Name: Thyroperoxidase, Inhibition
Key Event Component
| Process | Object | Action |
|---|---|---|
| iodide peroxidase activity | thyroid peroxidase | decreased |
AOPs Including This Key Event
Stressors
| Name |
|---|
| 2(3H)-Benzothiazolethione |
| 2-mercaptobenzothiazole |
| Ethylene thiourea |
| Mercaptobenzothiazole |
| Methimazole |
| Propylthiouracil |
| Resorcinol |
| Thiouracil |
| Ethylenethiourea |
| Amitrole |
| 131-55-5 |
| 2,2',4,4'-Tetrahydroxybenzophenone |
| Daidzein |
| Genistein |
| 4-Nonylphenol |
| 4-propoxyphenol |
| Sulfamethazine |
Biological Context
| Level of Biological Organization |
|---|
| Molecular |
Cell term
| Cell term |
|---|
| thyroid follicular cell |
Organ term
| Organ term |
|---|
| thyroid follicle |
Evidence for Perturbation by Stressor
Overview for Molecular Initiating Event
There is a wealth of information on the inhibition of TPO by drugs such as MMI and PTU, as well as environmental xenobiotics. In the landmark paper on thyroid disruption by environmental chemicals, Brucker-Davis (1998) identified environmental chemicals that depressed TH synthesis by inhibiting TPO. Hurley (1998) listed TPO as a major target for thyroid tumor inducing pesticides. More recent work has tested over 1000 chemicals using a high-throughput screening assay (Paul-Friedman et al., 2016).
Domain of Applicability
Taxonomic Applicability| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| rat | Rattus norvegicus | High | NCBI |
| humans | Homo sapiens | High | NCBI |
| pigs | Sus scrofa | High | NCBI |
| Xenopus laevis | Xenopus laevis | High | NCBI |
| chicken | Gallus gallus | High | NCBI |
| zebrafish | Danio rerio | High | NCBI |
| fathead minnow | Pimephales promelas | High | NCBI |
| mouse | Mus musculus | NCBI |
| Life Stage | Evidence |
|---|---|
| All life stages | High |
| Sex | Evidence |
|---|---|
| Female | High |
| Male | High |
Taxonomic: This KE is plausibly applicable across vertebrates. TPO inhibition is a MIE conserved across taxa, with supporting data from experimental models and human clinical testing. This conservation is likely a function of the high degree of protein sequence similarity in the catalytic domain of mammalian peroxidases (Taurog, 1999). Ample data available for human, rat, and porcine TPO inhibition demonstrate qualitative concordance across these species (Schmultzer et al., 2007; Paul et al., 2013; Hornung et al., 2010). A comparison of rat TPO and pig TPO, bovine lactoperoxidase, and human TPO inhibition by genistein demonstrated good qualitative and quantitative (40–66%) inhibition across species, as indicated by quantification of MIT and DIT production (Doerge and Chang, 2002). Ealey et al. (1984) demonstrated peroxidase activity in guinea pig thyroid tissue using 3,3'-diaminobenzidine tetrahydrochloride (DAB) as a substrate that is oxidized by the peroxidase to form a brown insoluble reaction product. Formation of this reaction product was inhibited by 3-amino-1,2,4-triazole and the TPO inhibitor, methimazole (MMI). A comparative analysis of this action of MMI between rat- and human-derived TPO indicates concordance of qualitative response. Data also suggest an increased quantitative sensitivity to MMI in rat compared to human (Vickers et al., 2012). Paul et al. (2013) tested 12 chemicals using the guaiacol assay using both porcine and rat thyroid microsomes. The authors concluded that there was an excellent qualitative concordance between rat and porcine TPO inhibition, as all chemicals that inhibited TPO in porcine thyroid microsomes also inhibited TPO in rat thyroid microsomes when tested within the same concentration range. In addition, these authors noted a qualitative concordance that ranged from 1.5 to 50-fold differences estimated by relative potency. Similary, Takayama et al. (1986) found a very large species difference in potency for sulfamonomethoxine between cynomologus monkeys and rats.
Life stage: Applicability to certain life stages may depend on the species and their dependence on maternally transferred thyroid hormones during the earliest phases of development. The earliest life stages of teleost fish rely on maternally transferred THs to regulate certain developmental processes until embryonic TH synthesis is active (Power et al., 2001). As a result, TPO inhibition is not expected to decrease TH synthesis during these earliest stages of development. In zebrafish, Opitz et al. (2011) showed the formation of a first thyroid follicle at 55 hours post fertilization (hpf), Chang et al. (2012) showed a first significant TH increase at 120 hpf and Walter et al. (2019) showed clear TH production already at 72 hpf and not at 24 hpf but did not analyse time points between 24 and 72 hpf. In fathead minnows, a significant increase of whole body thyroid hormone levels was already observed between 1 and 2 dpf, which corresponds to the appearance of the thyroid anlage at 35 hpf prior to the first observation of thyroid follicles at 58 hpf (Wabuke-Bunoti and Firling, 1983). It is still uncertain when exactly embryonic TH synthesis is activated and how this determines sensitivity to TPO inhibition.
Sex: This KE is plausibly applicable to both sexes. The molecular components responsible for thyroid hormone synthesis, including thyroperoxidase, are identical in both sexes. Therefore inhibition of deiodinases is not expected to be sex-specific.
Key Event Description
Thyroperoxidase (TPO) is a heme-containing apical membrane protein within the follicular lumen of thyrocytes that acts as the enzymatic catalyst for thyroid hormone (TH) synthesis. TPO catalyzes several reactions in the thyroid gland, including: the oxidation of iodide; nonspecific iodination of tyrosyl residues of thyroglobulin (Tg); and, the coupling of iodotyrosyls to produce Tg-bound monoiodotyrosine (MIT) and diiodotyrosine (DIT) (Divi et al., 1997; Kessler et al., 2008; Ruf et al., 2006; Taurog et al., 1996). The outcome of TPO inhibition is decreased synthesis of thyroxine (T4) and triiodothyronine (T3), a decrease in release of these hormones from the gland into circulation, and unless compensated, a consequent decrease in systemic concentrations of T4, and possibly T3. The primary product of TPO-catalyzed TH synthesis is T4 (Taurog et al., 1996; Zoeller et al., 2007) that would be peripherally or centrally deiodinated to T3.
It is important to note that TPO is a complex enzyme and that has two catalytic cycles and is capable of iodinating multiple species (Divi et al., 1997). Alterations in all of these events are not covered by some of the commonly used assays that measure “TPO inhibition” (e.g., guaiacol and AmplexUltraRed, see below). Therefore, in the context of this AOP we are using TPO inhibiton not in the classical sense, but instead to refer to the empirical data derived from the assays commonly used to investigate environmental chemicals.
Figure 1 below illustrates the enzymatic and nonenzymatic reactions mediated by TPO that result in the synthesis of thyroxine (T4) .

Inhibition of TPO can be reversible, with transient interaction between the enzyme and the chemical, or irreversible, whereby suicide substrates permanently inactivate the enzyme. Reversible and irreversible TPO inhibition may be determined by the chemical structure, may be concentration dependent, or may be influenced by other conditions, including the availability of iodine (Doerge and Chang, 2002).
The ontogeny of TPO has been determined using both direct and indirect evidence in mammals. Available evidence suggests the 11th to 12th fetal week as the beginning of functional TPO in humans. In rodents, TPO function begins late in the second fetal week, with the first evidence of T4 secretion on gestational day 17 (Remy et al., 1980). Thyroid-specific genes appear in the thyroid gland according to a specific temporal pattern; thyroglobulin (Tg), TPO (Tpo), and TSH receptor (Tshr) genes are expressed by gestational day 14 in rats, and the sodium iodide symporter, NIS (Nis), is expressed by gestational day 16 in rats. Maturation to adult function is thought to occur within a few weeks after parturition in rats and mice, and within the first few months in neonatal humans (Santisteban and Bernal, 2005). Tg is first detected in human fetuses starting at 5th week of gestation and rises throughout gestation (Thorpe-Beeston et al., 1992), but iodine trapping and T4 production does not occur until around 10-12 weeks. Also, the dimerization of Tg, a characteristic of adult TH storage, is not found until much later in human gestation (Pintar, 2000). In rats, Tg immunoreactivity does not appear until day 15 of gestation (Fukiishi et al., 1982; Brown et al., 2000). The vast majority of research and knowledge on Tg is from mammals, although genomic orthologs are known for a variety of other species (Holzer et al., 2016). It is important to note that prior to the onset of fetal thyroid function, TH are still required by the developing fetus which until that time relies solely on maternal sources. Chemical-induced TPO inhibition can affect synthesis in the maternal gland and in the fetal gland.
The components of the TH system responsible for TH synthesis are highly conserved across vertebrates. In fish and amphibians TPO and NIS inhibition result in an expected decrease of TH synthesis (Hornung et al., 2010; Tietge et al., 2013; Nelson et al., 2016; Stinckens et al., 2016; Stinckens et al., 2020) like in mammals. Although the thyroid hormone system is highly conserved across vertebrates, there are some taxon-specific considerations.
Zebrafish and fathead minnows are oviparous fish species in which maternal thyroid hormones are transferred to the eggs and regulate early embryonic developmental processes during external (versus intra-uterine in mammals) development (Power et al., 2001; Campinho et al., 2014; Ruuskanen and Hsu, 2018) until embryonic thyroid hormone synthesis is initiated. Maternal transfer of thyroid hormones to the eggs has been demonstrated in zebrafish (Walpita et al., 2007; Chang et al., 2012) and fathead minnows (Crane et al., 2004; Nelson et al., 2016).
Inhibition of thyroperoxidase can only occur after activation of embryonic TH synthesis mediated by thyroperoxidase. Endogenous transcription profiles of thyroid-related genes in zebrafish and fathead minnow showed that mRNA coding for thyroid peroxidase is maternally transferred in relatively high amounts with subsequent mRNA degradation followed by initiation of embryonic transcription around hatching (Vergauwen et al., 2018).
How it is Measured or Detected
There are no approved OECD or EPA guideline study protocols for measurement of TPO inhibition. However, there is an OECD scoping document on identification of chemicals that modulate TH signaling that provides details on a TPO assay (OECD, 2017).
From the early 1960's, microsomal fractions prepared from porcine thyroid glands and isolated porcine follicles were used as a source of TPO for inhibition experiments (Taurog, 2005). Microsomes from human goiter samples (Vickers et al., 2012) and rat thyroid glands (Paul et al., 2013; 2014; Paul-Friedman et al., 2016) have also been used as a source of TPO.
TPO activity has been measured for decades via indirect assessment by kinetic measurement of the oxidation of guaiacol (Chang & Doerge 2000; Hornung et al., 2010; Schmutzler et al., 2007). This method is a low-throughput assay due to the very rapid kinetics of the guaiacol oxidation reaction. More recently, higher-throughput methods using commercial fluorescent and luminescent substrates with rodent, porcine, and human microsomal TPO have been developed (Vickers et al., 2012; Paul et al., 2013; 2014; Kaczur et al., 1997). This assay substitutes a pre-fluorescent substrate (Amplex UltraRed) for guaiacol, that when incubated with a source of peroxidase and excess hydrogen peroxidase, results in a stable fluorescent product proportional to TPO activity (Vickers et al., 2012). The stability of the fluorescent reaction product allows this assay to be used in a higher throughput format (Paul-Friedman et al., 2016). This approach is appropriate for high-throughput screening but does not elucidate the specific mechanism by which a chemical may inhibit TPO (Paul-Friedman et al., 2016), and as with most in vitro assays, is subject to various sources of assay interference (Thorne et al., 2010).
HPLC has been used to measure the activity of TPO via formation of the precursors monoiodotyrosine (MIT), diiodotyrosine (DIT), and both T3 and T4, in a reaction mixture containing TPO, or a surrogate enzyme such as lactoperoxidase (Divi & Doerge 1994). The tools and reagents for this method are all available. However, HPLC or other analytical chemistry techniques make this a low throughput assay, depending on the level of automation. A primary advantage of this in vitro method is that it directly informs hypotheses regarding the specific mechanism by which a chemical may impact thyroid hormone synthesis in vitro.
In fish, increases of TPO mRNA levels are often used as indirect evidence of TPO inhibition in in vivo experiments (Baumann et al., 2016; Nelson et al., 2016; Wang et al., 2020).
References
Baumann L, Ros A, Rehberger K, Neuhauss SCF, Segner H. 2016. Thyroid disruption in zebrafish (danio rerio) larvae: Different molecular response patterns lead to impaired eye development and visual functions. Aquatic Toxicology. 172:44-55.
Brown RS, Shalhoub V, Coulter S, Alex S, Joris I, De Vito W, Lian J, Stein GS. Developmental regulation of thyrotropin receptor gene expression in the fetal and neonatal rat thyroid: relation to thyroid morphology and to thyroid-specific gene expression. Endocrinology. 2000 Jan;141(1):340-5.
Brucker-Davis F. 1998. Effects of environmental synthetic chemicals on thyroid function. Thyroid 8:827-856.
Campinho MA, Saraiva J, Florindo C, Power DM. 2014. Maternal thyroid hormones are essential for neural development in zebrafish. Molecular Endocrinology. 28(7):1136-1149.
Chang, H. C. and D. R. Doerge (2000) Dietary genistein inactivates rat thyroid peroxidase in vivo without an apparent hypothyroid effect. Toxicol Appl Pharmacol. 168:244-252.
Chang J, Wang M, Gui W, Zhao Y, Yu L, Zhu G. 2012. Changes in thyroid hormone levels during zebrafish development. Zoological Science. 29(3):181-184.
Crane HM, Pickford DB, Hutchinson TH, Brown JA. 2004. Developmental changes of thyroid hormones in the fathead minnow, pimephales promelas. General and Comparative Endocrinology. 139(1):55-60.
Divi, R. L., & Doerge, D. R. (1994). Mechanism-based inactivation of lactoperoxidase and thyroid peroxidase by resorcinol derivatives. Biochemistry 33(32), 9668–9674.
Divi, R. L., Chang, H. C., & Doerge, D. R. (1997). Anti-Thyroid Isoflavones from Soybean. Biochem. Pharmacol. 54(10), 1087–1096.
Doerge DR, Chang HC. Inactivation of thyroid peroxidase by soy isoflavones, in vitro and in vivo. J Chromatogr B Analy Technol Biomed Life Sci. 2002 Sep 25;777(1-2):269-79.
Ealey PA, Henderson B, Loveridge N.A quantitative study of peroxidase activity in unfixed tissue sections of the guinea-pig thyroid gland. Histochem J. 1984 Feb;16(2):111-22.
Fukiishi Y, Harauchi T, Yoshizaki T, Hasegawa Y, Eguchi Y. Ontogeny of thyroid peroxidase activity in perinatal rats. Acta Endocrinol (Copenh). 1982 101(3):397-402.
Holzer G, Morishita Y, Fini JB, Lorin T, Gillet B, Hughes S, Tohmé M, Deléage G, Demeneix B, Arvan P, Laudet V. Thyroglobulin Represents a Novel Molecular Architecture of Vertebrates. J Biol Chem. 2016 Jun 16.
Hornung, M. W., Degitz, S. J., Korte, L. M., Olson, J. M., Kosian, P. a, Linnum, A. L., & Tietge, J. E. (2010). Inhibition of thyroid hormone release from cultured amphibian thyroid glands by methimazole, 6-propylthiouracil, and perchlorate. Toxicol Sci 118(1), 42–51.
Hurley PM. 1998. Mode of carcinogenic action of pesticides inducing thyroid follicular cell tumors in rodents. Environ Health Perspect 106:437-445.
Kaczur, V., Vereb, G., Molnár, I., Krajczár, G., Kiss, E., Farid, N. R., & Balázs, C. (1997). Effect of anti-thyroid peroxidase (TPO) antibodies on TPO activity measured by chemiluminescence assay. Clin. Chem 43(8 Pt 1), 1392–6.
Kessler, J., Obinger, C., Eales, G., 2008. Factors influencing the study of peroxidase- generated iodine species and implications for thyroglobulin synthesis. Thyroid 18, 769–774.
Nelson K, Schroeder A, Ankley G, Blackwell B, Blanksma C, Degitz S, Flynn K, Jensen K, Johnson R, Kahl M et al. 2016. Impaired anterior swim bladder inflation following exposure to the thyroid peroxidase inhibitor 2-mercaptobenzothiazole part i: Fathead minnow. Aquatic Toxicology. 173:192-203.
OECD (2017) New Scoping Document on in vitro and ex vivo Assays for the Identification of Modulators of Thyroid Hormone Signalling. Series on Testing and Assessment. No. 207. ISSN: 20777876 (online) http://dx.doi.org/10.1787/20777876
Opitz R, Maquet E, Zoenen M, Dadhich R, Costagliola S. 2011. Tsh receptor function is required for normal thyroid differentiation in zebrafish. Molecular Endocrinology. 25(9):1579-1599.
Paul KB, Hedge JM, Macherla C, Filer DL, Burgess E, Simmons SO, Crofton KM, Hornung MW. Cross-species analysis of thyroperoxidase inhibition by xenobiotics demonstrates conservation of response between pig and rat. Toxicology. 2013. 312:97-107
Paul, K.B., Hedge, J.M., Rotroff, D.M., Hornung, M.W., Crofton, K.M., Simmons, S.O. 2014. Development of a thyroperoxidase inhibition assay for high-throughput screening. Chem. Res. Toxicol. 27(3), 387-399.
Paul-Friedman K, Watt ED, Hornung MW, Hedge JM, Judson RS, Crofton KM, Houck KA, Simmons SO. 2016. Tiered High-Throughput Screening Approach to Identify Thyroperoxidase Inhibitors Within the ToxCast Phase I and II Chemical Libraries. Toxicol Sci. 151:160-80.
Pintar, J.E. (2000) Normal development of the hypothalamic-pituitary-thyroid axis. In. Werner & Ingbar’s The Thyroid. (8th ed), Braverman. L.E. and Utiger, R.D. (eds) Lippincott Williams and Wilkins, Philadelphia.
Power DM, Llewellyn L, Faustino M, Nowell MA, Bjornsson BT, Einarsdottir IE, Canario AV, Sweeney GE. 2001. Thyroid hormones in growth and development of fish. Comp Biochem Physiol C Toxicol Pharmacol. 130(4):447-459.
Remy L, Michel-Bechet M, Athouel-Haon AM, Magre S. Critical study of endogenous peroxidase activity: its role in the morphofunctional setting of the thyroid follicle in the rat fetus. Acta Histochem. 1980;67(2):159-72.
Ruf, J., & Carayon, P. (2006). Structural and functional aspects of thyroid peroxidase. Archives of Biochemistry and Biophysics, 445(2), 269–77.
Ruuskanen S, Hsu BY. 2018. Maternal thyroid hormones: An unexplored mechanism underlying maternal effects in an ecological framework. Physiological and Biochemical Zoology. 91(3):904-916.
Santisteban P, Bernal J. Thyroid development and effect on the nervous system. Rev Endocr Metab Disord. 2005 Aug;6(3):217-28.
Schmutzler, C., Bacinski, A., Gotthardt, I., Huhne, K., Ambrugger, P., Klammer, H., Schlecht, C., Hoang-Vu, C., Gruters, A., Wuttke, W., Jarry, H., Kohrle, J., 2007a. The ultraviolet filter benzophenone 2 interferes with the thyroid hormone axis in rats and is a potent in vitro inhibitor of human recombinant thyroid peroxidase. Endocrinology 148, 2835–2844.
Stinckens E, Vergauwen L, Blackwell BR, Anldey GT, Villeneuve DL, Knapen D. 2020. Effect of thyroperoxidase and deiodinase inhibition on anterior swim bladder inflation in the zebrafish. Environmental Science & Technology. 54(10):6213-6223.
Stinckens E, Vergauwen L, Schroeder A, Maho W, Blackwell B, Witters H, Blust R, Ankley G, Covaci A, Villeneuve D et al. 2016. Impaired anterior swim bladder inflation following exposure to the thyroid peroxidase inhibitor 2-mercaptobenzothiazole part ii: Zebrafish. Aquatic Toxicology. 173:204-217.
Taurog A. 2005. Hormone synthesis. In: Werner and Ingbar’s The Thyroid: A Fundamental and Clinical Text (Braverman LE, Utiger RD, eds). Philadelphia:Lippincott, Williams and Wilkins, 47–81
Taurog, a, Dorris, M. L., & Doerge, D. R. (1996). Mechanism of simultaneous iodination and coupling catalyzed by thyroid peroxidase. Archives of Biochemistry and Biophysics, Taurog A. Molecular evolution of thyroid peroxidase. Biochimie. 1999 May;81(5):557-62
Takayama S, Aihara K, Onodera T, Akimoto T. Antithyroid effects of propylthiouracil and sulfamonomethoxine in rats and monkeys. Toxicol Appl Pharmacol. 1986 Feb;82(2):191-9.
Thorne N, Auld DS, Inglese J. Apparent activity in high-throughput screening: origins of compound-dependent assay interference. Curr Opin Chem Biol. 2010 Jun;14(3):315-24.
Thorpe-Beeston JG, Nicolaides KH, McGregor AM. Fetal thyroid function. Thyroid. 1992 Fall;2(3):207-17. Review.
Tietge JE, Degitz SJ, Haselman JT, Butterworth BC, Korte JJ, Kosian PA, Lindberg-Livingston AJ, Burgess EM, Blackshear PE, Hornung MW. 2013. Inhibition of the thyroid hormone pathway in xenopus laevis by 2-mercaptobenzothiazole. Aquatic Toxicology. 126:128-136.
Vergauwen L, Cavallin JE, Ankley GT, Bars C, Gabriels IJ, Michiels EDG, Fitzpatrick KR, Periz-Stanacev J, Randolph EC, Robinson SL et al. 2018. Gene transcription ontogeny of hypothalamic-pituitary-thyroid axis development in early-life stage fathead minnow and zebrafish. General and Comparative Endocrinology. 266:87-100.
Vickers AE, Heale J, Sinclair JR, Morris S, Rowe JM, Fisher RL. Thyroid organotypic rat and human cultures used to investigate drug effects on thyroid function, hormone synthesis and release pathways. Toxicol Appl Pharmacol. 2012 Apr 1;260(1):81-8.
Wabukebunoti MAN, Firling CE. 1983. The prehatching development of the thyroid-gland of the fathead minnow, pimephales-promelas (rafinesque). General and Comparative Endocrinology. 49(2):320-331.
Walpita CN, Van der Geyten S, Rurangwa E, Darras VM. 2007. The effect of 3,5,3'-triiodothyronine supplementation on zebrafish (danio rerio) embryonic development and expression of iodothyronine deiodinases and thyroid hormone receptors. Gen Comp Endocrinol. 152(2-3):206-214.
Walter KM, Miller GW, Chen XP, Yaghoobi B, Puschner B, Lein PJ. 2019. Effects of thyroid hormone disruption on the ontogenetic expression of thyroid hormone signaling genes in developing zebrafish (danio rerio). General and Comparative Endocrinology. 272:20-32.
Wang JX, Shi GH, Yao JZ, Sheng N, Cui RN, Su ZB, Guo Y, Dai JY. 2020. Perfluoropolyether carboxylic acids (novel alternatives to pfoa) impair zebrafish posterior swim bladder development via thyroid hormone disruption. Environment International. 134.
Zoeller, R. T., Tan, S. W., & Tyl, R. W. (2007). General background on the hypothalamic-pituitary-thyroid (HPT) axis. Critical Reviews in Toxicology, 37(1-2), 11–53.
List of Key Events in the AOP
Event: 277: Thyroid hormone synthesis, Decreased
Short Name: TH synthesis, Decreased
Key Event Component
| Process | Object | Action |
|---|---|---|
| thyroid hormone generation | thyroid hormone | decreased |
AOPs Including This Key Event
Stressors
| Name |
|---|
| Propylthiouracil |
| Methimazole |
Biological Context
| Level of Biological Organization |
|---|
| Cellular |
Cell term
| Cell term |
|---|
| thyroid follicular cell |
Organ term
| Organ term |
|---|
| thyroid gland |
Evidence for Perturbation by Stressor
Propylthiouracil
6-n-proylthiouracil is a common positive control for inhibition of TPO
Methimazole
Methimazole is a very common positve control for inhibition of TPO
Domain of Applicability
Taxonomic Applicability| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| rat | Rattus norvegicus | High | NCBI |
| human | Homo sapiens | High | NCBI |
| Xenopus laevis | Xenopus laevis | Moderate | NCBI |
| zebrafish | Danio rerio | High | NCBI |
| fathead minnow | Pimephales promelas | Moderate | NCBI |
| Sus scrofa | Sus scrofa | High | NCBI |
| Life Stage | Evidence |
|---|---|
| All life stages | High |
| Sex | Evidence |
|---|---|
| Male | High |
| Female | High |
Taxonomic: This KE is plausibly applicable across vertebrates. Decreased TH synthesis resulting from TPO or NIS inhibition is conserved across vertebrate taxa, with in vivo evidence from humans, rats, amphibians, some fish specis, and birds, and in vitro evidence from rat and porcine microsomes. Indeed, TPO and NIS mutations result in congenital hypothyroidism in humans (Bakker et al., 2000; Spitzweg and Morris, 2010), demonstrating the essentiality of TPO and NIS function toward maintaining euthyroid status. Though decreased serum T4 is used as a surrogate measure to indicate chemical-mediated decreases in TH synthesis, clinical and veterinary management of hyperthyroidism and Grave's disease using propylthiouracil and methimazole, known to decrease TH synthesis, indicates strong medical evidence for chemical inhibition of TPO (Zoeller and Crofton, 2005).
Life stage: Applicability to certain life stages may depend on the species and their dependence on maternally transferred thyroid hormones during the earliest phases of development. The earliest life stages of teleost fish (e.g., fathead minnow, zebrafish) rely on maternally transferred THs to regulate certain developmental processes until embryonic TH synthesis is active (Power et al., 2001). In externally developing fish species, decreases in TH synthesis can only occur after initiation of embryonic TH synthesis. In zebrafish, Opitz et al. (2011) showed the formation of a first thyroid follicle at 55 hours post fertilization (hpf), Chang et al. (2012) showed a first significant TH increase at 120 hpf and Walter et al. (2019) showed clear TH production already at 72 hpf but did not analyse time points between 24 and 72 hpf. Therefore, it is still uncertain when exactly embryonic TH synthesis is activated and thus when exactly this process becomes sensitive to disruption. In fathead minnows, a significant increase of whole body thyroid hormone levels was already observed between 1 and 2 dpf, which corresponds to the appearance of the thyroid anlage at 35 hpf prior to the first observation of thyroid follicles at 58 hpf (Wabuke-Bunoti and Firling, 1983). It currently remains unclear when exactly embryonic thyroid hormone production is initiated in zebrafish.
Sex: The KE is plausibly applicable to both sexes. Thyroid hormones are essential in both sexes and the components of the HPT-axis are identical in both sexes. There can however be sex-dependent differences in the sensitivity to the disruption of thyroid hormone levels and the magnitude of the response. In humans, females appear more susceptible to hypothyroidism compared to males when exposed to certain halogenated chemicals (Hernandez‐Mariano et al., 2017; Webster et al., 2014). In adult zebrafish, Liu et al. (2019) showed sex-dependent changes in thyroid hormone levels and mRNA expression of regulatory genes including corticotropin releasing hormone (crh), thyroid stimulating hormone (tsh) and deiodinase 2 after exposure to organophosphate flame retardants. The underlying mechanism of any sex-related differences remains unclear.
Key Event Description
The thyroid hormones (TH), triiodothyronine (T3) and thyroxine (T4) are thyrosine based hormones. Synthesis of TH is regulated by thyroid-stimulating hormone (TSH) binding to its receptor and thyroidal availability of iodine via the sodium iodide symporter (NIS). Other proteins contributing to TH production in the thyroid gland, including thyroperoxidase (TPO), dual oxidase enzymes (DUOX), and pendrin are also necessary for iodothyronine production (Zoeller et al., 2007).
The production of THs in the thyroid gland and resulting serum concentrations are controlled by a negatively regulated feedback mechanism. Decreased T4 and T3 serum concentrations activates the hypothalamus-pituitary-thyroid (HPT) axis which upregulates thyroid-stimulating hormone (TSH) that acts to increase production of additional THs (Zoeller and Tan, 2007). This regulatory system includes: 1) the hypothalamic secretion of the thyrotropin-releasing hormone (TRH); 2) the thyroid-stimulating hormone (TSH) secretion from the anterior pituitary; 3) hormonal transport by the plasma binding proteins; 4) cellular uptake mechanisms at the tissue level; 5) intracellular control of TH concentration by deiodinating mechanisms; 6) transcriptional function of the nuclear TH receptor; and 7) in the fetus, the transplacental passage of T4 and T3 (Zoeller et al., 2007).
TRH and the TSH primarily regulate the production of T4, often considered a “pro-hormone,” and to a lesser extent of T3, the transcriptionally active TH. Most of the hormone released from the thyroid gland into circulation is in the form of T4, while peripheral deiodination of T4 is responsible for the majority of circulating T3. Outer ring deiodination of T4 to T3 is catalyzed by the deiodinases 1 and 2 (DIO1 and DIO2), with DIO1 expressed mainly in liver and kidney, and DIO2 expressed in several tissues including the brain (Bianco et al., 2006). Conversion of T4 to T3 takes place mainly in liver and kidney, but also in other target organs such as in the brain, the anterior pituitary, brown adipose tissue, thyroid and skeletal muscle (Gereben et al., 2008; Larsen, 2009).
In mammals, most evidence for the ontogeny of TH synthesis comes from measurements of serum hormone concentrations. And, importantly, the impact of xenobiotics on fetal hormones must include the influence of the maternal compartment since a majority of fetal THs are derived from maternal blood early in fetal life, with a transition during mid-late gestation to fetal production of THs that is still supplemented by maternal THs. In humans, THs can be found in the fetus as early as gestational weeks 10-12, and concentations rise continuously until birth. At term, fetal T4 is similar to maternal levels, but T3 remains 2-3 fold lower than maternal levels. In rats, THs can be detected in the fetus as early as the second gestational week, but fetal synthesis does not start until gestational day 17 with birth at gestational day 22-23. Maternal THs continue to supplement fetal production until parturition. (see Howdeshell, 2002; Santisteban and Bernal, 2005 for review). The ontogeny of TPO inhibition during development by environmental chemicals is a data gap.
Decreased TH synthesis in the thyroid gland may result from several possible molecular-initiating events (MIEs) including: 1) Disruption of key catalytic enzymes or cofactors needed for TH synthesis, including TPO, NIS, or dietary iodine insufficiency. Theoretically, decreased synthesis of Tg could also affect TH production (Kessler et al., 2008; Yi et al., 1997). Mutations in genes that encode requisite proteins in the thyroid may also lead to impaired TH synthesis, including mutations in pendrin associated with Pendred Syndrome (Dossena et al., 2011), mutations in TPO and Tg (Huang and Jap 2015), and mutations in NIS (Spitzweg and Morris, 2010). 2) Decreased TH synthesis in cases of clinical hypothyroidism may be due to Hashimoto's thyroiditis or other forms of thyroiditis, or physical destruction of the thyroid gland as in radioablation or surgical treatment of thyroid lymphoma. 3) It is possible that TH synthesis may also be reduced subsequent to disruption of the negative feedback mechanism governing TH homeostasis, e.g. pituitary gland dysfunction may result in a decreased TSH signal with concomitant T3 and T4 decreases. 4) More rarely, hypothalamic dysfunction can result in decreased TH synthesis.
Increased fetal thyroid levels are also possible. Maternal Graves disease, which results in fetal thyrotoxicosis (hyperthyroidism and increased serum T4 levels), has been successfully treated by maternal administration of TPO inhibitors (c.f., Sato et al., 2014).
It should be noted that different species and different lifestages store different amounts of TH precursor and iodine within the thyroid gland. Thus, decreased TH synthesis via transient iodine insufficiency or inhibition of TPO may not affect TH release from the thyroid gland until depletion of stored iodinated Tg. Adult humans may store sufficient Tg-DIT residues to serve for several months to a year of TH demand (Greer et al., 2002; Zoeller, 2004). Neonates and infants have a much more limited supply of less than a week.
While the thyroid hormone system is highly conserved across vertebrates, there are some taxon-specific considerations.
Zebrafish and fathead minnows are oviparous fish species in which maternal thyroid hormones are transferred to the eggs and regulate early embryonic developmental processes during external (versus intra-uterine in mammals) development (Power et al., 2001; Campinho et al., 2014; Ruuskanen and Hsu, 2018) until embryonic thyroid hormone synthesis is initiated. Maternal transfer of thyroid hormones to the eggs has been demonstrated in zebrafish (Walpita et al., 2007; Chang et al., 2012) and fathead minnows (Crane et al., 2004; Nelson et al., 2016).
Decreases in TH synthesis can only occur after initiation of embryonic TH synthesis. The components of the TH system responsible for TH synthesis are highly conserved across vertebrates and therefore interference with the same molecular targets compared to mammals can lead to decreased TH synthesis (TPO, NIS, etc.) in fish. Endogenous transcription profiles of thyroid-related genes in zebrafish and fathead minnow showed that mRNA coding for these genes is also maternally transferred and increasing expression of most transcripts during hatching and embryo-larval transition indicates a fully functional HPT axis in larvae (Vergauwen et al., 2018). Although the HPT axis is highly conserved, there are some differences between fish and mammals (Blanton and Specker, 2007; Deal and Volkoff, 2020). For example, in fish, corticotropin releasing hormone (CRH) often plays a more important role in regulating thyrotropin (TSH) secretion by the pituitary and thus thyroid hormone synthesis compared to TSH-releasing hormone (TRH). Also, in most fish species thyroid follicles are more diffusely located in the pharyngeal region rather than encapsulated in a gland.
How it is Measured or Detected
Decreased TH synthesis is often implied by measurement of TPO and NIS inhibition measured clinically and in laboratory models as these enzymes are essential for TH synthesis. Rarely is decreased TH synthesis measured directly, but rather the impact of chemicals on the quantity of T4 produced in the thyroid gland, or the amount of T4 present in serum is used as a marker of decreased T4 release from the thyroid gland (e.g., Romaldini et al., 1988). Methods used to assess TH synthesis include, incorporation of radiolabel tracer compounds, radioimmunoassay, ELISA, and analytical detection.
Recently, amphibian thyroid explant cultures have been used to demonstrate direct effects of chemicals on TH synthesis, as this model contains all necessary synthesis enzymes including TPO and NIS (Hornung et al., 2010). For this work THs was measured by HPLC/ICP-mass spectometry. Decreased TH synthesis and release, using T4 release as the endpoint, has been shown for thiouracil antihyperthyroidism drugs including MMI, PTU, and the NIS inhibitor perchlorate (Hornung et al., 2010).
Techniques for in vivo analysis of thyroid hormone system disruption among other drug-related effects in fish were reviewed by Raldua and Piña (2014). TIQDT (Thyroxine-immunofluorescence quantitative disruption test) is a method that provides an immunofluorescent based estimate of thyroxine in the gland of zebrafish (Raldua and Babin, 2009; Thienpont et al., 2011; Jomaa et al., 2014; Rehberger et al., 2018). Thienpont used this method with ~25 xenobiotics (e.g., amitrole, perchlorate, methimazole, PTU, DDT, PCBs). The method detected changes for all chemicals known to directly impact TH synthesis in the thyroid gland (e.g., NIS and TPO inbibitors), but not those that upregulate hepatic catabolism of T4. Rehberger et al. (2018) updated the method to enable simultaneous semi-quantitative visualization of intrafollicular T3 and T4 levels. Most often, whole body thyroid hormone level measurements in fish early life stages are used as indirect evidence of decreased thyroid hormone synthesis (Nelson et al., 2016; Stinckens et al., 2016; Stinckens et al., 2020). Analytical determination of thyroid hormone levels by LC-MS is becoming increasingly available (Hornung et al., 2015).
More recently, transgenic zebrafish with fluorescent thyroid follicles are being used to visualize the compensatory proliferation of the thyroid follicles following inhibition of thyroid hormone synthesis (Opitz et al., 2012).
References
Bakker B, Bikker H, Vulsma T, de Randamie JS, Wiedijk BM, De Vijlder JJ. 2000. Two decades of screening for congenital hypothyroidism in The Netherlands: TPO gene mutations in total iodide organification defects (an update). The Journal of clinical endocrinology and metabolism. 85:3708-3712.
Bianco AC, Kim BW. (2006). Deiodinases: implications of the local control of thyroid hormone action. J Clin Invest. 116: 2571–2579.
Blanton ML, Specker JL. 2007. The hypothalamic-pituitary-thyroid (hpt) axis in fish and its role in fish development and reproduction. Crit Rev Toxicol. 37(1-2):97-115.
Campinho MA, Saraiva J, Florindo C, Power DM. 2014. Maternal thyroid hormones are essential for neural development in zebrafish. Molecular Endocrinology. 28(7):1136-1149.
Chang J, Wang M, Gui W, Zhao Y, Yu L, Zhu G. 2012. Changes in thyroid hormone levels during zebrafish development. Zoological Science. 29(3):181-184.
Crane HM, Pickford DB, Hutchinson TH, Brown JA. 2004. Developmental changes of thyroid hormones in the fathead minnow, pimephales promelas. General and Comparative Endocrinology. 139(1):55-60.
Deal CK, Volkoff H. 2020. The role of the thyroid axis in fish. Frontiers in Endocrinology. 11.
Dossena S, Nofziger C, Brownstein Z, Kanaan M, Avraham KB, Paulmichl M. (2011). Functional characterization of pendrin mutations found in the Israeli and Palestinian populations. Cell Physiol Biochem. 28: 477-484.Gereben B, Zavacki AM, Ribich S, Kim BW, Huang SA, Simonides WS, Zeöld A, Bianco AC. (2008). Cellular and molecular basis of deiodinase-regulated thyroid hormone signalling. Endocr Rev. 29:898–938.
Gereben B, Zeöld A, Dentice M, Salvatore D, Bianco AC. Activation and inactivation of thyroid hormone by deiodinases: local action with general consequences. Cell Mol Life Sci. 2008 Feb;65(4):570-90
Greer MA, Goodman G, Pleus RC, Greer SE. Health effects assessment for environmental perchlorate contamination: the dose response for inhibition of thyroidal radioiodine uptake in humans. Environ Health Perspect. 2002. 110:927-937.
Hernandez-Mariano JA, Torres-Sanchez L, Bassol-Mayagoitia S, Escamilla-Nunez M, Cebrian ME, Villeda-Gutierrez EA, Lopez-Rodriguez G, Felix-Arellano EE, Blanco-Munoz J. 2017. Effect of exposure to p,p '-dde during the first half of pregnancy in the maternal thyroid profile of female residents in a mexican floriculture area. Environmental Research. 156:597-604.
Hornung MW, Degitz SJ, Korte LM, Olson JM, Kosian PA, Linnum AL, Tietge JE. 2010. Inhibition of thyroid hormone release from cultured amphibian thyroid glands by methimazole, 6-propylthiouracil, and perchlorate. Toxicol Sci 118:42-51.
Hornung MW, Kosian PA, Haselman JT, Korte JJ, Challis K, Macherla C, Nevalainen E, Degitz SJ. 2015. In vitro, ex vivo, and in vivo determination of thyroid hormone modulating activity of benzothiazoles. Toxicological Sciences. 146(2):254-264.
Howdeshell KL. 2002. A model of the development of the brain as a construct of the thyroid system. Environ Health Perspect. 110 Suppl 3:337-48.
Huang CJ and Jap TS. 2015. A systematic review of genetic studies of thyroid disorders in Taiwan. J Chin Med Assoc. 78: 145-153.
Jomaa B, Hermsen SAB, Kessels MY, van den Berg JHJ, Peijnenburg AACM, Aarts JMMJG, Piersma AH, Rietjens IMCM. 2014. Developmental toxicity of thyroid-active compounds in a zebrafish embryotoxicity test. Altex-Alternatives to Animal Experimentation. 31(3):303-317.
Kessler J, Obinger C, Eales G. Factors influencing the study of peroxidase-generated iodine species and implications for thyroglobulin synthesis. Thyroid. 2008 Jul;18(7):769-74. doi: 10.1089/thy.2007.0310
Larsen PR. (2009). Type 2 iodothyronine deiodinase in human skeletal muscle: new insights into its physiological role and regulation. J Clin Endocrinol Metab. 94:1893-1895.
Liu XS, Cai Y, Wang Y, Xu SH, Ji K, Choi K. 2019. Effects of tris(1,3-dichloro-2-propyl) phosphate (tdcpp) and triphenyl phosphate (tpp) on sex-dependent alterations of thyroid hormones in adult zebrafish. Ecotoxicology and Environmental Safety. 170:25-32.
Nelson K, Schroeder A, Ankley G, Blackwell B, Blanksma C, Degitz S, Flynn K, Jensen K, Johnson R, Kahl M et al. 2016. Impaired anterior swim bladder inflation following exposure to the thyroid peroxidase inhibitor 2-mercaptobenzothiazole part i: Fathead minnow. Aquatic Toxicology. 173:192-203.
Opitz R, Maquet E, Huisken J, Antonica F, Trubiroha A, Pottier G, Janssens V, Costagliola S. 2012. Transgenic zebrafish illuminate the dynamics of thyroid morphogenesis and its relationship to cardiovascular development. Developmental Biology. 372(2):203-216.
Opitz R, Maquet E, Zoenen M, Dadhich R, Costagliola S. 2011. Tsh receptor function is required for normal thyroid differentiation in zebrafish. Molecular Endocrinology. 25(9):1579-1599.
Power DM, Llewellyn L, Faustino M, Nowell MA, Bjornsson BT, Einarsdottir IE, Canario AV, Sweeney GE. 2001. Thyroid hormones in growth and development of fish. Comp Biochem Physiol C Toxicol Pharmacol. 130(4):447-459.
Raldua D, Babin PJ. 2009. Simple, rapid zebrafish larva bioassay for assessing the potential of chemical pollutants and drugs to disrupt thyroid gland function. Environmental Science & Technology. 43(17):6844-6850.
Raldua D, Pina B. 2014. In vivo zebrafish assays for analyzing drug toxicity. Expert Opinion on Drug Metabolism & Toxicology. 10(5):685-697.
Rehberger K, Baumann L, Hecker M, Braunbeck T. 2018. Intrafollicular thyroid hormone staining in whole-mount zebrafish (danio rerio) embryos for the detection of thyroid hormone synthesis disruption. Fish Physiology and Biochemistry. 44(3):997-1010.
Romaldini JH, Farah CS, Werner RS, Dall'Antonia Júnior RP, Camargo RS. 1988. "In vitro" study on release of cyclic AMP and thyroid hormone in autonomously functioning thyroid nodules. Horm Metab Res.20:510-2.
Ruuskanen S, Hsu BY. 2018. Maternal thyroid hormones: An unexplored mechanism underlying maternal effects in an ecological framework. Physiological and Biochemical Zoology. 91(3):904-916.
Santisteban P, Bernal J. Thyroid development and effect on the nervous system. Rev Endocr Metab Disord. 2005 Aug;6(3):217-28.
Spitzweg C, Morris JC. 2010. Genetics and phenomics of hypothyroidism and goiter due to NIS mutations. Molecular and cellular endocrinology. 322:56-63.
Stinckens E, Vergauwen L, Blackwell BR, Anldey GT, Villeneuve DL, Knapen D. 2020. Effect of thyroperoxidase and deiodinase inhibition on anterior swim bladder inflation in the zebrafish. Environmental Science & Technology. 54(10):6213-6223.
Stinckens E, Vergauwen L, Schroeder A, Maho W, Blackwell B, Witters H, Blust R, Ankley G, Covaci A, Villeneuve D et al. 2016. Impaired anterior swim bladder inflation following exposure to the thyroid peroxidase inhibitor 2-mercaptobenzothiazole part ii: Zebrafish. Aquatic Toxicology. 173:204-217.
Thienpont B, Tingaud-Sequeira A, Prats E, Barata C, Babin PJ, Raldúa D. Zebrafish eleutheroembryos provide a suitable vertebrate model for screening chemicals that impair thyroid hormone synthesis. Environ Sci Technol. 2011. 45(17):7525-32.
Vergauwen L, Cavallin JE, Ankley GT, Bars C, Gabriels IJ, Michiels EDG, Fitzpatrick KR, Periz-Stanacev J, Randolph EC, Robinson SL et al. 2018. Gene transcription ontogeny of hypothalamic-pituitary-thyroid axis development in early-life stage fathead minnow and zebrafish. General and Comparative Endocrinology. 266:87-100.
Wabukebunoti MAN, Firling CE. 1983. The prehatching development of the thyroid-gland of the fathead minnow, pimephales-promelas (rafinesque). General and Comparative Endocrinology. 49(2):320-331.
Walpita CN, Van der Geyten S, Rurangwa E, Darras VM. 2007. The effect of 3,5,3'-triiodothyronine supplementation on zebrafish (danio rerio) embryonic development and expression of iodothyronine deiodinases and thyroid hormone receptors. Gen Comp Endocrinol. 152(2-3):206-214.
Walter KM, Miller GW, Chen XP, Yaghoobi B, Puschner B, Lein PJ. 2019. Effects of thyroid hormone disruption on the ontogenetic expression of thyroid hormone signaling genes in developing zebrafish (danio rerio). General and Comparative Endocrinology. 272:20-32.
Webster GM, Venners SA, Mattman A, Martin JW. 2014. Associations between perfluoroalkyl acids (pfass) and maternal thyroid hormones in early pregnancy: A population-based cohort study. Environmental Research. 133:338-347.
Yi X, Yamamoto K, Shu L, Katoh R, Kawaoi A. Effects of Propyithiouracil (PTU) Administration on the Synthesis and Secretion of Thyroglobulin in the Rat Thyroid Gland: A Quantitative Immuno-electron Microscopic Study Using Immunogold Technique. Endocr Pathol. 1997 Winter;8(4):315-325.
Zoeller RT, Crofton KM. 2005. Mode of action: developmental thyroid hormone insufficiency--neurological abnormalities resulting from exposure to propylthiouracil. Crit Rev Toxicol. 35:771-81
Zoeller RT, Tan SW, Tyl RW. 2007. General background on the hypothalamic-pituitary-thyroid (HPT) axis. Critical reviews in toxicology. 37:11-53.
Zoeller RT. Interspecies differences in susceptibility to perturbation of thyroid hormone homeostasis requires a definition of "sensitivity" that is informative for risk analysis. Regul Toxicol Pharmacol. 2004 Dec;40(3):380.
Event: 281: Thyroxine (T4) in serum, Decreased
Short Name: T4 in serum, Decreased
Key Event Component
| Process | Object | Action |
|---|---|---|
| abnormal circulating thyroxine level | thyroxine | decreased |
AOPs Including This Key Event
Stressors
| Name |
|---|
| Propylthiouracil |
| Methimazole |
| Perchlorate |
Biological Context
| Level of Biological Organization |
|---|
| Tissue |
Organ term
| Organ term |
|---|
| serum |
Evidence for Perturbation by Stressor
Propylthiouracil
6-n-propylthouracil is a classic positive control for inhibition of TPO
Methimazole
Methimazole is a classic positive control for inhibition of TPO.
Perchlorate
Perchlorate ion (ClO− ₄) is a classic positive control for inhibition of NIS
Domain of Applicability
Taxonomic Applicability| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| human | Homo sapiens | High | NCBI |
| rat | Rattus norvegicus | High | NCBI |
| mouse | Mus musculus | High | NCBI |
| chicken | Gallus gallus | Moderate | NCBI |
| Xenopus laevis | Xenopus laevis | Moderate | NCBI |
| zebrafish | Danio rerio | High | NCBI |
| fathead minnow | Pimephales promelas | High | NCBI |
| Sus scrofa | Sus scrofa | High | NCBI |
| Life Stage | Evidence |
|---|---|
| All life stages | High |
| Sex | Evidence |
|---|---|
| Female | High |
| Male | High |
Taxonomic: This KE is plausibly applicable across vertebrates and the overall evidence supporting taxonomic applicability is strong. THs are evolutionarily conserved molecules present in all vertebrate species (Hulbert, 2000; Yen, 2001). Moreover, their crucial role in zebrafish development, embryo-to-larval transition and larval-to-juvenile transition (Thienpont et al., 2011; Liu and Chan, 2002), and amphibian and lamprey metamorphoses is well established (Manzon and Youson, 1997; Yaoita and Brown, 1990; Furlow and Neff, 2006). Their existence and importance has also been described in many different animal and plant kingdoms (Eales, 1997; Heyland and Moroz, 2005), while their role as environmental messenger via exogenous routes in echinoderms confirms the hypothesis that these molecules are widely distributed among the living organisms (Heyland and Hodin, 2004). However, the role of TH in the different species depends on the expression and function of specific proteins (e.g receptors or enzymes) under TH control and may vary across species and tissues. As such extrapolation regarding TH action across species and developmental stages should be done with caution.
With few exceptions, vertebrate species have circulating T4 (and T3) that are bound to transport proteins in blood. Clear species differences exist in serum transport proteins (Dohler et al., 1979; Yamauchi and Isihara, 2009). There are three major transport proteins in mammals; thyroid binding globulin (TBG), transthyretin (TTR), and albumin. In adult humans, the percent bound to these proteins is about 75, 15 and 10 percent, respectively (Schussler 2000). In contrast, in adult rats the majority of THs are bound to TTR. Thyroid binding proteins are developmentally regulated in rats. TBG is expressed in rats until approximately postnatal day (PND) 60, with peak expression occurring during weaning (Savu et al., 1989). However, low levels of TBG persist into adult ages in rats and can be experimentally induced by hypothyroidism, malnutrition, or caloric restriction (Rouaze-Romet et al., 1992). While these species differences impact TH half-life (Capen, 1997) and possibly regulatory feedback mechanisms, there is little information on quantitative dose-response relationships of binding proteins and serum hormones during development across different species. Serum THs are still regarded as the most robust measurable key event causally linked to downstream adverse outcomes.
Life stage: The earliest life stages of teleost fish rely on maternally transferred THs to regulate certain developmental processes until embryonic TH synthesis is active (Power et al., 2001). As a result, T4 levels are not expected to decrease in response to exposure to inhibitors of TH synthesis during these earliest stages of development. In zebrafish, Opitz et al. (2011) showed the formation of a first thyroid follicle at 55 hours post fertilization (hpf), Chang et al. (2012) showed a first significant TH increase at 120 hpf and Walter et al. (2019) showed clear TH production already at 72 hpf but did not analyse time points between 24 and 72 hpf. In fathead minnows, a significant increase of whole body thyroid hormone levels was already observed between 1 and 2 dpf, which corresponds to the appearance of the thyroid anlage at 35 hpf prior to the first observation of thyroid follicles at 58 hpf (Wabuke-Bunoti and Firling, 1983). It is still uncertain when exactly embryonic TH synthesis is activated and how this determines sensitivity to TH disruptors.
Sex: The KE is plausibly applicable to both sexes. Thyroid hormones are essential in both sexes and the components of the HPT-axis are identical in both sexes. There can however be sex-dependent differences in the sensitivity to the disruption of thyroid hormone levels and the magnitude of the response. In humans, females appear more susceptible to hypothyroidism compared to males when exposed to certain halogenated chemicals (Hernandez‐Mariano et al., 2017; Webster et al., 2014). In adult zebrafish, Liu et al. (2019) showed sex-dependent changes in thyroid hormone levels and mRNA expression of regulatory genes including corticotropin releasing hormone (crh), thyroid stimulating hormone (tsh) and deiodinase 2 after exposure to organophosphate flame retardants. The underlying mechanism of any sex-related differences remains unclear.
Key Event Description
All iodothyronines are derived from the modification of tyrosine molecules (Taurog, 2000). There are two biologically active thyroid hormones (THs) in serum, triiodothyronine (T3) and T4, and a few less active iodothyronines (rT3, 3,5-T2). T4 is the predominant TH in circulation, comprising approximately 80% of the TH excreted from the thyroid gland in mammals and is the pool from which the majority of T3 in serum is generated (Zoeller et al., 2007). As such, serum T4 changes usually precede changes in other serum THs. Decreased thyroxine (T4) in serum results from one or more MIEs upstream and is considered a key biomarker of altered TH homeostasis (DeVito et al., 1999).
Serum T4 is used as a biomarker of TH status because the circulatory system serves as the major transport and delivery system for TH delivery to tissues. The majority of THs in the blood are bound to transport proteins (Bartalena and Robbins, 1993). In serum, it is the unbound, or ‘free’ form of the hormone that is thought to be available for transport into tissues. Free hormones are approximately 0.03 and 0.3 percent for T4 and T3, respectively. There are major species differences in the predominant binding proteins and their affinities for THs (see below). However, there is broad agreement that changes in serum concentrations of THs is diagnostic of thyroid disease or chemical-induced disruption of thyroid homeostasis across vertebrates (DeVito et al., 1999; Miller et al., 2009; Zoeller et al., 2007; Carr and Patiño, 2011).
Normal serum T4 reference ranges can be species and lifestage specific. In rodents, serum THs are low in the fetal circulation, increasing as the fetal thyroid gland becomes functional on gestational day 17, just a few days prior to birth. After birth serum hormones increase steadily, peaking at two weeks, and falling slightly to adult levels by postnatal day 21 (Walker et al., 1980; Harris et al., 1978; Goldey et al., 1995; Lau et al., 2003). Similarly, in humans, adult reference ranges for THs do not reflect the normal ranges for children at different developmental stages, with TH concentrations highest in infants, still increased in childhood, prior to a decline to adult levels coincident with pubertal development (Corcoran et al. 1977; Kapelari et al., 2008).
In some frog species, there is an analogous peak in thyroid hormones in tadpoles that starts around embryonic NF stage 56, peaks at Stage 62 and the declines to lower levels by Stage 56 (Sternberg et al., 2011; Leloup and Buscaglia, 1977).
Additionally, ample evidence is available from studies investigating responses to inhibitors of thyroid hormone synthesis in fish. For example, Stinckens et al. (2020) showed reduced whole body T4 concentrations in zebrafish larvae exposed to 50 or 100 mg/L methimazole, a potent TPO inhibitor, from immediately after fertilization until 21 or 32 days of age. Exposure to 37 or 111 mg/L propylthiouracil also reduced T4 levels after exposure up to 14, 21 and 32 days in the same study. Walter et al. (2019) showed that propylthiouracil had no effect on T4 levels in 24h old zebrafish, but decreased T4 levels of 72h old zebrafish. This difference is probably due to the onset of embryonic TH production between the age of 24 and 72 hours (Opitz et al., 2011). Stinckens et al. (2016) showed that exposure to 2-mercaptobenzothiazole (MBT), an environmentally relevant TPO inhibitor, decreased whole body T4 levels in continuously exposed 5 and 32 day old zebrafish larvae. A high concentration of MBT also decreased whole body T4 levels in 6 day old fathead minnows, but recovery was observed at the age of 21 days although the fish were kept in the exposure medium (Nelson et al., 2016). Crane et al. (2006) showed decreased T4 levels in 28 day old fathead minnows continuously exposed to 32 or 100 µg/L methimazole.
How it is Measured or Detected
Serum T3 and T4 can be measured as free (unbound) or total (bound + unbound). Free hormone concentrations are clinically considered more direct indicators of T4 and T3 activities in the body, but in animal studies, total T3 and T4 are typically measured. Historically, the most widely used method in toxicology is the radioimmunoassay (RIA). The method is routinely used in rodent endocrine and toxicity studies. The ELISA method is commonly used as a human clinical test method. Analytical determination of iodothyronines (T3, T4, rT3, T2) and their conjugates, through methods employing HPLC, liquid chromatography, immuno luminescence, and mass spectrometry are less common, but are becoming increasingly available (Hornung et al., 2015; DeVito et al., 1999; Baret and Fert, 1989; Spencer, 2013; Samanidou V.F et al., 2000; Rathmann D. et al., 2015 ). In fish early life stages most evidence for the ontogeny of thyroid hormone synthesis comes from measurements of whole body thyroid hormone levels using LC-MS techniques (Hornung et al., 2015) are increasingly used to accurately quantify whole body thyroid hormone levels as a proxy for serum thyroid hormone levels (Nelson et al., 2016; Stinckens et al., 2016; Stinckens et al., 2020). It is important to note that thyroid hormones concentrations can be influenced by a number of intrinsic and extrinsic factors (e.g., circadian rhythms, stress, food intake, housing, noise) (see for example, Döhler et al., 1979).
Any of these measurements should be evaluated for the relationship to the actual endpoint of interest, repeatability, reproducibility, and lower limits of quantification using a fit-for-purpose approach (i.e., different regulatory needs will require different levels of confidence in the AOP). This is of particular significance when assessing the very low levels of TH present in fetal serum. Detection limits of the assay must be compatible with the levels in the biological sample. All three of the methods summarized above would be fit-for-purpose, depending on the number of samples to be evaluated and the associated costs of each method. Both RIA and ELISA measure THs by an indirect methodology, whereas analytical determination is the most direct measurement available. All these methods, particularly RIA, are repeatable and reproducible.
References
Axelrad DA, Baetcke K, Dockins C, Griffiths CW, Hill RN, Murphy PA, Owens N, Simon NB, Teuschler LK. Risk assessment for benefits analysis: framework for analysis of a thyroid-disrupting chemical. J Toxicol Environ Health A. 2005 68(11-12):837-55.
Baret A. and Fert V. T4 and ultrasensitive TSH immunoassays using luminescent enhanced xanthine oxidase assay. J Biolumin Chemilumin. 1989. 4(1):149-153
Bartalena L, Robbins J. Thyroid hormone transport proteins. Clin Lab Med. 1993 Sep;13(3):583-98. Bassett JH, Harvey CB, Williams GR. (2003). Mechanisms of thyroid hormone receptor-specific nuclear and extra nuclear actions. Mol Cell Endocrinol. 213:1-11.
Capen CC. Mechanistic data and risk assessment of selected toxic end points of the thyroid gland. Toxicol Pathol. 1997 25(1):39-48.
Carr JA, Patino R. 2011. The hypothalamus-pituitary-thyroid axis in teleosts and amphibians: Endocrine disruption and its consequences to natural populations. General and Comparative Endocrinology. 170(2):299-312.
Chang J, Wang M, Gui W, Zhao Y, Yu L, Zhu G. 2012. Changes in thyroid hormone levels during zebrafish development. Zoological Science. 29(3):181-184.
Cope RB, Kacew S, Dourson M. A reproductive, developmental and neurobehavioral study following oral exposure of tetrabromobisphenol A on Sprague-Dawley rats. Toxicology. 2015 329:49-59.
Corcoran JM, Eastman CJ, Carter JN, Lazarus L. (1977). Circulating thyroid hormone levels in children. Arch Dis Child. 52: 716-720.
Crane HM, Pickford DB, Hutchinson TH, Brown JA. 2006. The effects of methimazole on development of the fathead minnow, pimephales promelas, from embryo to adult. Toxicological Sciences. 93(2):278-285.
Crofton KM. Developmental disruption of thyroid hormone: correlations with hearing dysfunction in rats. Risk Anal. 2004 Dec;24(6):1665-71.
DeVito M, Biegel L, Brouwer A, Brown S, Brucker-Davis F, Cheek AO, Christensen R, Colborn T, Cooke P, Crissman J, Crofton K, Doerge D, Gray E, Hauser P, Hurley P, Kohn M, Lazar J, McMaster S, McClain M, McConnell E, Meier C, Miller R, Tietge J, Tyl R. (1999). Screening methods for thyroid hormone disruptors. Environ Health Perspect. 107:407-415.
Döhler KD, Wong CC, von zur Mühlen A (1979). The rat as model for the study of drug effects on thyroid function: consideration of methodological problems. Pharmacol Ther B. 5:305-18.
Eales JG. (1997). Iodine metabolism and thyroid related functions in organisms lacking thyroid follicles: Are thyroid hormones also vitaminsProc Soc Exp Biol Med. 214:302-317.
Furlow JD, Neff ES. (2006). A developmental switch induced by thyroid hormone: Xenopus laevis metamorphosis. Trends Endocrinol Metab. 17:40–47.
Goldey ES, Crofton KM. Thyroxine replacement attenuates hypothyroxinemia, hearing loss, and motor deficits following developmental exposure to Aroclor 1254 in rats. Toxicol Sci. 1998 45(1):94-10
Goldey ES, Kehn LS, Lau C, Rehnberg GL, Crofton KM. Developmental exposure to polychlorinated biphenyls (Aroclor 1254) reduces circulating thyroid hormone concentrations and causes hearing deficits in rats. Tox Appl Pharmacol. 1995 135(1):77-88.
Harris AR, Fang SL, Prosky J, Braverman LE, Vagenakis AG. Decreased outer ring monodeiodination of thyroxine and reverse triiodothyronine in the fetal and neonatal rat. Endocrinology. 1978 Dec;103(6):2216-22
Hernandez-Mariano JA, Torres-Sanchez L, Bassol-Mayagoitia S, Escamilla-Nunez M, Cebrian ME, Villeda-Gutierrez EA, Lopez-Rodriguez G, Felix-Arellano EE, Blanco-Munoz J. 2017. Effect of exposure to p,p '-dde during the first half of pregnancy in the maternal thyroid profile of female residents in a mexican floriculture area. Environmental Research. 156:597-604.
Heyland A, Hodin J. (2004). Heterochronic developmental shift caused by thyroid hormone in larval sand dollars and its implications for phenotypic plasticity and the evolution of non-feeding development. Evolution. 58: 524-538.
Heyland A, Moroz LL. (2005). Cross-kingdom hormonal signaling: an insight from thyroid hormone functions in marine larvae. J Exp Biol. 208:4355-4361.
Hill RN, Crisp TM, Hurley PM, Rosenthal SL, Singh DV. Risk assessment of thyroid follicular cell tumors. Environ Health Perspect. 1998 Aug;106(8):447-57.
Hornung MW, Kosian P, Haselman J, Korte J, Challis K, Macherla C, Nevalainen E, Degitz S (2015) In vitro, ex vivo and in vivo determination of thyroid hormone modulating activity of benzothiazoles. Toxicol Sci 146:254-264.
Hulbert AJ. Thyroid hormones and their effects: a new perspective. Biol Rev Camb Philos Soc. 2000 Nov;75(4):519-631. Review.
Kapelari K, Kirchlechner C, Högler W, Schweitzer K, Virgolini I, Moncayo R. 2008. Pediatric reference intervals for thyroid hormone levels from birth to adulthood: a retrospective study. BMC Endocr Disord. 8: 15.
Lau C, Thibodeaux JR, Hanson RG, Rogers JM, Grey BE, Stanton ME, Butenhoff JL, Stevenson LA. Exposure to perfluorooctane sulfonate during pregnancy in rat and mouse. II: postnatal evaluation. Toxicol Sci. 2003 Aug;74(2):382-92.
Leloup, J., and M. Buscaglia. La triiodothyronine: hormone de la métamorphose des amphibiens. CR Acad Sci 284 (1977): 2261-2263.
Liu J, Liu Y, Barter RA, Klaassen CD.: Alteration of thyroid homeostasis by UDP-glucuronosyltransferase inducers in rats: a dose-response study. J Pharmacol Exp Ther 273, 977-85, 1994
Liu XS, Cai Y, Wang Y, Xu SH, Ji K, Choi K. 2019. Effects of tris(1,3-dichloro-2-propyl) phosphate (tdcpp) and triphenyl phosphate (tpp) on sex-dependent alterations of thyroid hormones in adult zebrafish. Ecotoxicology and Environmental Safety. 170:25-32.
Liu YW, Chan WK. 2002. Thyroid hormones are important for embryonic to larval transitory phase in zebrafish. Differentiation. 70(1):36-45.
Manzon RG, Youson JH. (1997). The effects of exogenous thyroxine (T4) or triiodothyronine (T3), in the presence and absence of potassium perchlorate, on the incidence of metamorphosis and on serum T4 and T3 concentrations in larval sea lampreys (Petromyzon marinus L.). Gen Comp Endocrinol. 106:211-220.
McClain RM. Mechanistic considerations for the relevance of animal data on thyroid neoplasia to human risk assessment. Mutat Res. 1995 Dec;333(1-2):131-42
Miller MD, Crofton KM, Rice DC, Zoeller RT. Thyroid-disrupting chemicals: interpreting upstream biomarkers of adverse outcomes. Environ Health Perspect. 2009 117(7):1033-41
Morse DC, Wehler EK, Wesseling W, Koeman JH, Brouwer A. Alterations in rat brain thyroid hormone status following pre- and postnatal exposure to polychlorinated biphenyls (Aroclor 1254). Toxicol Appl Pharmacol. 1996 Feb;136(2):269-79.
Nelson K, Schroeder A, Ankley G, Blackwell B, Blanksma C, Degitz S, Flynn K, Jensen K, Johnson R, Kahl M et al. 2016. Impaired anterior swim bladder inflation following exposure to the thyroid peroxidase inhibitor 2-mercaptobenzothiazole part i: Fathead minnow. Aquatic Toxicology. 173:192-203.
NTP National Toxicology Program.: NTP toxicology and carcinogenesis studies of 3,3'-dimethylbenzidine dihydrochloride (CAS no. 612-82-8) in F344/N rats (drinking water studies). Natl Toxicol Program Tech Rep Ser 390, 1-238, 1991.
O'Connor, J. C., J. C. Cook, et al. (1998). "An ongoing validation of a Tier I screening battery for detecting endocrine-active compounds (EACs)." Toxicol Sci 46(1): 45-60.
O'Connor, J. C., L. G. Davis, et al. (2000). "Detection of dopaminergic modulators in a tier I screening battery for identifying endocrine-active compounds (EACs)." Reprod Toxicol 14(3): 193-205.
Opitz R, Maquet E, Zoenen M, Dadhich R, Costagliola S. 2011. Tsh receptor function is required for normal thyroid differentiation in zebrafish. Molecular Endocrinology. 25(9):1579-1599.
Power DM, Llewellyn L, Faustino M, Nowell MA, Bjornsson BT, Einarsdottir IE, Canario AV, Sweeney GE. 2001. Thyroid hormones in growth and development of fish. Comp Biochem Physiol C Toxicol Pharmacol. 130(4):447-459.
Rathmann D, Rijntjes E, Lietzow J, Köhrle J. (2015) Quantitative Analysis of Thyroid Hormone Metabolites in Cell Culture Samples Using LC-MS/MS. Eur Thyroid J. Sep;4(Suppl 1):51-8.
Rouaze-Romet M, Savu L, Vranckx R, Bleiberg-Daniel F, Le Moullac B, Gouache P, Nunez EA. 1992. Reexpression of thyroxine-binding globulin in postweaning rats during protein or energy malnutrition. Acta Endocrinol (Copenh).127:441-448.
Samanidou VF, Kourti PV. (2009) Rapid HPLC method for the simultaneous monitoring of duloxetine, venlaflaxine, fluoxetine and paroxetine in biofluids. Bioanalysis. 2009 Aug;1(5):905-17.
Savu L, Vranckx R, Maya M, Gripois D, Blouquit MF, Nunez EA. 1989. Thyroxine-binding globulin and thyroxinebinding prealbumin in hypothyroid and hyperthyroid developing rats. BiochimBiophys Acta. 992:379-384.
Schneider S, Kaufmann W, Strauss V, van Ravenzwaay B. Vinclozolin: a feasibility and sensitivity study of the ILSI-HESI F1-extended one-generation rat reproduction protocol. Regul Toxicol Pharmacol. 2011 Feb;59(1):91-100.
Schussler, G.C. (2000). The thyroxine-binding proteins. Thyroid 10:141–149.
Spencer, CA. (2013). Assay of thyroid hormone and related substances. In De Groot, LJ et al. (Eds). Endotext. South Dartmouth, MA
Sternberg RM, Thoemke KR, Korte JJ, Moen SM, Olson JM, Korte L, Tietge JE, Degitz SJ Jr. Control of pituitary thyroid-stimulating hormone synthesis and secretion by thyroid hormones during Xenopus metamorphosis. Gen Comp Endocrinol. 2011. 173(3):428-37
Stinckens E, Vergauwen L, Blackwell BR, Anldey GT, Villeneuve DL, Knapen D. 2020. Effect of thyroperoxidase and deiodinase inhibition on anterior swim bladder inflation in the zebrafish. Environmental Science & Technology. 54(10):6213-6223.
Stinckens E, Vergauwen L, Schroeder A, Maho W, Blackwell B, Witters H, Blust R, Ankley G, Covaci A, Villeneuve D et al. 2016. Impaired anterior swim bladder inflation following exposure to the thyroid peroxidase inhibitor 2-mercaptobenzothiazole part ii: Zebrafish. Aquatic Toxicology. 173:204-217.
Taurog A. 2005. Hormone synthesis. In: Werner and Ingbar’s The Thyroid: A Fundamental and Clinical Text (Braverman LE, Utiger RD, eds). Philadelphia:Lippincott, Williams and Wilkins, 47–81Walker P, Dubois JD, Dussault JH. Free thyroid hormone concentrations during postnatal development in the rat. Pediatr Res. 1980 Mar;14(3):247-9.
Thienpont B, Tingaud-Sequeira A, Prats E, Barata C, Babin PJ, Raldúa D. Zebrafish eleutheroembryos provide a suitable vertebrate model for screening chemicals that impair thyroid hormone synthesis. Environ Sci Technol. 2011 Sep 1;45(17):7525-32.
Wabukebunoti MAN, Firling CE. 1983. The prehatching development of the thyroid-gland of the fathead minnow, pimephales-promelas (rafinesque). General and Comparative Endocrinology. 49(2):320-331.
Walter KM, Miller GW, Chen XP, Yaghoobi B, Puschner B, Lein PJ. 2019. Effects of thyroid hormone disruption on the ontogenetic expression of thyroid hormone signaling genes in developing zebrafish (danio rerio). General and Comparative Endocrinology. 272:20-32.
Webster GM, Venners SA, Mattman A, Martin JW. 2014. Associations between perfluoroalkyl acids (pfass) and maternal thyroid hormones in early pregnancy: A population-based cohort study. Environmental Research. 133:338-347.
Yamauchi K1, Ishihara A. Evolutionary changes to transthyretin: developmentally regulated and tissue-specific gene expression. FEBS J. 2009. 276(19):5357-66.
Yaoita Y, Brown DD. (1990). A correlation of thyroid hormone receptor gene expression with amphibian metamorphosis. Genes Dev. 4:1917-1924.
Yen PM. (2001). Physiological and molecular basis of thyroid hormone action. Physiol Rev. 81:1097-1142.
Zoeller RT, Tan SW, Tyl RW. General background on the hypothalamic-pituitary-thyroid (HPT) axis. Crit Rev Toxicol. 2007 Jan-Feb;37(1-2):11-53
Zoeller, R. T., R. Bansal, et al. (2005). "Bisphenol-A, an environmental contaminant that acts as a thyroid hormone receptor antagonist in vitro, increases serum thyroxine, and alters RC3/neurogranin expression in the developing rat brain." Endocrinology 146(2): 607-612.
Event: 1003: Decreased, Triiodothyronine (T3) in serum
Short Name: Decreased, Triiodothyronine (T3) in serum
Key Event Component
| Process | Object | Action |
|---|---|---|
| abnormal circulating hormone level | 3,3',5'-triiodothyronine | decreased |
AOPs Including This Key Event
Biological Context
| Level of Biological Organization |
|---|
| Tissue |
Organ term
| Organ term |
|---|
| serum |
Domain of Applicability
Taxonomic Applicability| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| zebrafish | Danio rerio | High | NCBI |
| fathead minnow | Pimephales promelas | High | NCBI |
| African clawed frog | Xenopus laevis | High | NCBI |
| Life Stage | Evidence |
|---|---|
| All life stages | High |
| Sex | Evidence |
|---|---|
| Unspecific | Moderate |
Taxonomic: The overall evidence supporting taxonomic applicability is strong. With few exceptions vertebrate species have circulating T3 and T4 that are bound to transport proteins in blood. Therefore, the current key event is plausibly applicable to vertebrates in general. Clear species differences exist in transport proteins (Yamauchi and Isihara, 2009). Specifically, the majority of supporting data for TH decreases in serum come from rat studies, and the predominant iodothyronine binding protein in rat serum is transthyretin (TTR). TTR demonstrates a reduced binding affinity for T4 when compared with thyroxine binding globulin (TBG), the predominant serum binding protein for T4 in humans. This difference in serum binding protein affinity for THs is thought to modulate serum half-life for T4; the half-life of T4 in rats is 12-24 hr, wherease the half-life in humans is 5-9 days (Capen, 1997). While these species differences impact hormone half-life, possibly regulatory feedback mechanisms, and quantitative dose-response relationships, measurement of serum THs is still regarded as a measurable key event causatively linked to downstream adverse outcomes.
THs are evolutionarily conserved molecules present in all vertebrate species (Hulbert, 2000; Yen, 2001). Moreover, their crucial role in amphibian and larbean metamorphoses (Manzon and Youson, 1997; Yaoita and Brown, 1990) as well as fish development, embryo-to-larval transition and larval-to-juvenile transition (Thienpont et al., 2011; Liu and Chan, 2002) is well established. Their existence and importance has been also described in many differrent animal and plant kingdoms (Eales, 1997; Heyland and Moroz, 2005), while their role as environmental messenger via exogenous routes in echinoderms confirms the hypothesis that these molecules are widely distributed among the living organisms (Heyland and Hodin, 2004). However, the role of TH in the different species may differ depending on the expression or function of specific proteins (e.g receptors or enzymes) that are related to TH function, and therefore extrapolation between species should be done with caution.
Life stage: Thyroid hormones are essential in all life stages, but decreases of circulating levels are associated with specific developmental events. The earliest life stages of teleost fish rely on maternally transferred THs to regulate certain developmental processes until embryonic TH synthesis is active (Power et al., 2001). As a result, T4 levels are not expected to decrease in response to exposure to inhibitors of TH synthesis during these earliest stages of development. However, T3 levels are expected to decrease upon exposure to deiodinase inhibitors in any life stage, since maternal T4 needs to be activated to T3 by deiodinases similar to embryonically synthesized T4.
Sex: The KE is plausibly applicable to both sexes. Thyroid hormones are essential in both sexes and the components of the HPT-axis are identical in both sexes. There can however be sex-dependent differences in the sensitivity to the disruption of thyroid hormone levels and the magnitude of the response. In humans, females appear more susceptible to hypothyroidism compared to males when exposed to certain halogenated chemicals (Hernandez‐Mariano et al., 2017; Webster et al., 2014). In adult zebrafish, Liu et al. (2019) showed sex-dependent changes in thyroid hormone levels and mRNA expression of regulatory genes including corticotropin releasing hormone (crh), thyroid stimulating hormone (tsh) and deiodinase 2 after exposure to organophosphate flame retardants. The underlying mechanism of any sex-related differences remains unclear.
Key Event Description
There are two biologically active thyroid hormones (THs), triiodothyronine (T3) and thyroxine (T4), and a few less active iodothyronines (rT3, 3,5-T2), which are all derived from the modification of tyrosine molecules (Hulbert, 2000). However, the plasma concentrations of the other iodothyronines are significantly lower than those of T3 and T4. The different iodothyronines are formed by the sequential outer or inner ring monodeiodination of T4 by the deiodinating enzymes, Dio1, Dio2, and Dio3 (Gereben et al., 2008). Deiodinase structure is considered to be unique, as THs are the only molecules in the body that incorporate iodide.
The circulatory system serves as the major transport and delivery system for THs from synthesis in the gland to delivery to tissues. The majority of THs in the blood are bound to transport proteins (Bartalena and Robbins, 1993). In humans, the major transport proteins are TBG (thyroxine binding globulin), TTR (transthyretin) and albumin. The percent bound to these proteins in adult humans is about 75, 15 and 10 percent, respectively (Schussler 2000). Unbound (free) hormones are approximately 0.03 and 0.3 percent for T4 and T3, respectively. In serum, it is the free form of the hormone that is active.
There are major species differences in the predominant binding proteins and their affinities for THs (see section below on Taxonomic applicability). However, there is broad agreement that changes in serum concentrations of THs is diagnostic of thyroid disease or chemical-induced disruption of thyroid homeostasis (Zoeller et al., 2007).
It is notable that the changes measured in the TH concentration reflect mainly the changes in the serum transport proteins rather than changes in the thyroid status. These thyroid-binding proteins serve as hormonal store which ensure their even and constant distribution in the different tissues, while they protect the most sensitive ones in the case of severe changes in thyroid availability, like in thyroidectomies (Obregon et al., 1981). Until recently, it was believed that all of the effects of TH were mediated by the binding of T3 to the thyroid nuclear receptors (TRa and TRb), a notion which is now questionable due to the increasing evidence that support the non-genomic action of TH (Davis et al., 2010, Moeller et al., 2006). Many non-nuclear TH binding sites have been identified to date and they usually lead to rapid cellular response in TH-effects (Bassett et al., 2003), but the specific pathways that are activated in this regard need to be elucidated.
The production of THs in the thyroid gland and the circulation levels in the bloodstream are self-controlled by an efficiently regulated feedback mechanism across the Hypothalamus-Pituitary-Thyroid (HPT) axis. TH levels are regulated, not only in the plasma level, but also in the individual cell level, to maintain homeostasis. This is succeeded by the efficient regulatory mechanism of the thyroid hormone axis which consists of the following: (1) the hypothalamic secretion of the thyrotropin-releasing hormone (TRH), (2) the thyroid-stimulating hormone (TSH) secretion from the anterior pituitary, (3) hormonal transport by the plasma binding proteins, (4) cellular uptake mechanisms in the cell level, (5) intracellular control of TH concentration by the deiodinating mechanism (6) transcriptional function of the nuclear thyroid hormone receptor and (7) in the fetus, the transplacental passage of T4 and T3 (Cheng et al., 2010).
In regards to the brain, the TH concentration involves also an additional level of regulation, namely the hormonal transport through the Blood Brain Barrier (BBB) (Williams, 2008). The TRH and the TSH are actually regulating the production of pro-hormone T4 and in a lesser extent of T3, which is the biologically active TH. The rest of the required amount of T3 is produced by outer ring deiodination of T4 by the deiodinating enzymes D1 and D2 (Bianco et al., 2006), a process which takes place mainly in liver and kidneys but also in other target organs such as in the brain, the anterior pituitary, brown adipose tissue, thyroid and skeletal muscle (Gereben et al., 2008; Larsen, 2009). Both hormones exert their action in almost all tissues of mammals and they are acting intracellularly, and thus the uptake of T3 and T4 by the target cells is a crucial step of the overall pathway. The trans-membrane transport of TH is performed mainly through transporters that differ depending on the cell type (Hennemann et al., 2001; Friesema et al., 2005; Visser et al., 2008). Many transporter proteins have been identified up to date but the monocarboxylate transporters (Mct8, Mct10) and the anion-transporting polypeptide (OATP1c1) show the highest degree of affinity towards TH (Jansen et al., 2005).
T3 and T4 have significant effects on normal development, neural differentiation, growth rate and metabolism (Yen, 2001; Brent, 2012; Williams, 2008), with the most prominent ones to occur during the fetal development and early childhood. The clinical features of hypothyroidism and hyperthyroidism emphasize the pleiotropic effects of these hormones on many different pathways and target organs. The thyroidal actions though are not only restricted to mammals, as their high significance has been identified also for other vertebrates, with the most well-studied to be the amphibian metamorphosis (Furlow and Neff, 2006). The importance of the thyroid-regulated pathways becomes more apparent in iodine deficient areas of the world, where a higher rate of cretinism and growth retardation has been observed and linked to decreased TH levels (Gilbert et al., 2012). Another very common cause of severe hypothyroidism in human is the congenital hypothyroidism, but the manifestation of these effects is only detectable in the lack of adequate treatment and is mainly related to neurological impairment and growth retardation (Glinoer, 2001), emphasizing the role of TH in neurodevelopment in all above cases. In adults, the thyroid-related effects are mainly linked to metabolic activities, such as deficiencies in oxygen consumption, and in the metabolism of the vitamin, proteins, lipids and carbohydrates, but these defects are subtle and reversible (Oetting and Yen, 2007). Blood tests to detect the amount of thyroid hormone (T4) and thyroid stimulating hormone (TSH) are routinely done for newborn babies for the diagnosis of congenital hypothyroidism at the earliest stage possible.
Although the components of the thyroid hormone system as well as thyroid hormone synthesis and action are highly conserved across vertebrates, there are some taxon-specific considerations.
Although the HPT axis is highly conserved, there are some differences between fish and mammals (Blanton and Specker, 2007; Deal and Volkoff, 2020). For example, in fish, corticotropin releasing hormone (CRH) often plays a more important role in regulating thyrotropin (TSH) secretion by the pituitary and thus thyroid hormone synthesis compared to TSH-releasing hormone (TRH). TTRs from fish have low sequence identity with human TTR, for example seabream TTR has 54% sequence identity with human TTR but the only amino acid difference within the thyroxine-binding site is the conservative substitution of Ser117 in human TTR to Thr117 in seabream TTR (Santos and Power, 1999; Yamauchi et al., 1999; Eneqvist et al., 2004). In vitro binding experiments showed that TH disrupting chemicals bind with equal or weaker affinity to seabream TTR than to the human TTR with polar TH disrupting chemicals, in particular, showing a more than 500-fold lower affinity for seabream TTR compared to human TTR (Zhang et al., 2018).
Zebrafish and fathead minnows are oviparous fish species in which maternal thyroid hormones are transferred to the eggs and regulate early embryonic developmental processes during external (versus intra-uterine in mammals) development (Power et al., 2001; Campinho et al., 2014; Ruuskanen and Hsu, 2018) until embryonic thyroid hormone synthesis is initiated. Maternal transfer of thyroid hormones, both T4 and T3, to the eggs has been demonstrated in zebrafish (Walpita et al., 2007; Chang et al., 2012) and fathead minnows (Crane et al., 2004; Nelson et al., 2016).
Several studies have reported evidence of T3 decreases after exposure to TPO inhibitors and deiodinase inhibitors in early life stages of zebrafish (Stinckens et al., 2016; Stinckens et al., 2020; Wang et al., 2020) and fathead minnow (Nelson et al., 2016; Cavallin et al., 2017).
How it is Measured or Detected
T3 and T4 can be measured as free (unbound) or total (bound + unbound). Free hormone are considered more direct indicators of T4 and T3 activities in the body. The majority of T3 and T4 measurements are made using either RIA or ELISA kits. In animal studies, total T3 and T4 are typically measured as the concentrations of free hormone are very low and difficult to detect. Historically, the most widely used method in toxicology is RIA. The method is routinely used in rodent endocrine and toxicity studies. The ELISA method has become more routine in rodent studies. The ELISA method is a commonly used as a human clinical test method.
Recently, analytical determination of iodothyronines (T3, T4, rT3, T2) and their conjugates through methods employing HPLC and mass spectrometry have become more common (DeVito et al., 1999; Miller et al., 2009; Hornung et al., 2015; Nelson et al., 2016; Stinckens et al., 2016).
Any of these measurements should be evaluated for fit-for-purpose, relationship to the actual endpoint of interest, repeatability, and reproducibility. All three of the methods summarized above would be fit-for-purpose, depending on the number of samples to be evaluated and the associated costs of each method. Both RIA and ELISA measure THs by a an indirect methodology, whereas analytical determination is the most direct measurement available. All of these methods, particularly RIA, are repeatable and reproducible.
In fish early life stages most evidence for the ontogeny of thyroid hormone synthesis comes from measurements of whole body thyroid hormone levels and using LC-MS techniques (Hornung et al., 2015) are increasingly used to accurately quantify whole body thyroid hormone levels as a proxy for serum thyroid hormone levels (Nelson et al., 2016; Stinckens et al., 2016; Stinckens et al., 2020).
References
Bartalena L, Robbins J.Thyroid hormone transport proteins.Clin Lab Med. 1993 Sep;13(3):583-98.
Bassett JH, Harvey CB, Williams GR. (2003). Mechanisms of thyroid hormone receptor-specific nuclear and extra nuclear actions. Mol Cell Endocrinol. 213:1-11.
Bianco AC, Kim BW. (2006). Deiodinases: implications of the local control of thyroid hormone action. J Clin Invest. 116: 2571–2579.
Blanton ML, Specker JL. 2007. The hypothalamic-pituitary-thyroid (hpt) axis in fish and its role in fish development and reproduction. Crit Rev Toxicol. 37(1-2):97-115.
Brent GA. (2012). Mechanisms of thyroid hormone action. J Clin Invest. 122: 3035-3043.
Campinho MA, Saraiva J, Florindo C, Power DM. 2014. Maternal thyroid hormones are essential for neural development in zebrafish. Molecular Endocrinology. 28(7):1136-1149.
Cavallin JE, Ankley GT, Blackwell BR, Blanksma CA, Fay KA, Jensen KM, Kahl MD, Knapen D, Kosian PA, Poole ST et al. 2017. Impaired swim bladder inflation in early life stage fathead minnows exposed to a deiodinase inhibitor, iopanoic acid. Environmental Toxicology and Chemistry. 36(11):2942-2952.
Chang J, Wang M, Gui W, Zhao Y, Yu L, Zhu G. 2012. Changes in thyroid hormone levels during zebrafish development. Zoological Science. 29(3):181-184.
Cheng SY, Leonard JL, Davis PJ. (2010).Molecular aspects of thyroid hormone actions. Endocr Rev. 31:139–170.
Crane HM, Pickford DB, Hutchinson TH, Brown JA. 2004. Developmental changes of thyroid hormones in the fathead minnow, pimephales promelas. General and Comparative Endocrinology. 139(1):55-60.
Davis PJ, Zhou M, Davis FB, Lansing L, Mousa SA, Lin HY. (2010). Mini-review: Cell surface receptor for thyroid hormone and nongenomic regulation of ion fluxes in excitable cells. Physiol Behav. 99:237–239.
Deal CK, Volkoff H. 2020. The role of the thyroid axis in fish. Frontiers in Endocrinology. 11.
DeVito M, Biegel L, Brouwer A, Brown S, Brucker-Davis F, Cheek AO, Christensen R, Colborn T, Cooke P, Crissman J, Crofton K, Doerge D, Gray E, Hauser P, Hurley P, Kohn M, Lazar J, McMaster S, McClain M, McConnell E, *Meier C, Miller R, Tietge J, Tyl R. (1999). Screening methods for thyroid hormone disruptors. Environ Health Perspect. 107:407-415.
Eales JG. (1997). Iodine metabolism and thyroid related functions in organisms lacking thyroid follicles: Are thyroid hormones also vitamins? Proc Soc Exp Biol Med. 214:302-317.
Eneqvist T, Lundberg E, Karlsson A, Huang SH, Santos CRA, Power DM, Sauer-Eriksson AE. 2004. High resolution crystal structures of piscine transthyretin reveal different binding modes for triiodothyronine and thyroxine. Journal of Biological Chemistry. 279(25):26411-26416.
Friesema EC, Jansen J, Milici C, Visser TJ. (2005). Thyroid hormone transporters. Vitam Horm. 70: 137–167.
Furlow JD, Neff ES. (2006). A developmental switch induced by thyroid hormone: Xenopus laevis metamorphosis. Trends Endocrinol Metab. 17:40–47.
Gereben B, Zavacki AM, Ribich S, Kim BW, Huang SA, Simonides WS, Zeöld A, Bianco AC. (2008). Cellular and molecular basis of deiodinase-regulated thyroid hormone signalling. Endocr Rev. 29:898–938.
Gilbert ME, Rovet J, Chen Z, Koibuchi N. (2012).Developmental thyroid hormone disruption: prevalence, environmental contaminants and neurodevelopmental consequences. Neurotoxicology. 33: 842-852.
Glinoer D. (2001).Potential consequences of maternal hypothyroidism on the offspring: evidence and implications. Horm Res. 55:109-114.
Hennemann G, Docter R, Friesema EC, de Jong M, Krenning EP, Visser TJ. (2001). Plasma membrane transport of thyroid hormones and its role in thyroid hormone metabolism and bioavailability. Endocr Rev. 22:451-476.
Hernandez-Mariano JA, Torres-Sanchez L, Bassol-Mayagoitia S, Escamilla-Nunez M, Cebrian ME, Villeda-Gutierrez EA, Lopez-Rodriguez G, Felix-Arellano EE, Blanco-Munoz J. 2017. Effect of exposure to p,p '-dde during the first half of pregnancy in the maternal thyroid profile of female residents in a mexican floriculture area. Environmental Research. 156:597-604.
Heyland A, Hodin J. (2004). Heterochronic developmental shift caused by thyroid hormone in larval sand dollars and its implications for phenotypic plasticity and the evolution of non-feeding development. Evolution. 58: 524-538.
Heyland A, Moroz LL. (2005). Cross-kingdom hormonal signaling: an insight from thyroid hormone functions in marine larvae. J Exp Biol. 208:4355-4361.
Hornung, M.W., Kosian, P.A., Haselman, J.T., Korte, J.J., Challis, K., Macherla, C., Nevalainen, E., Degitz, S.J., 2015. In Vitro, Ex Vivo, and In Vivo Determination of Thyroid Hormone Modulating Activity of Benzothiazoles. Toxicological Sciences 146, 254-264.
Hulbert A J. (2000). Thyroid hormones and their effects: A new perspective. Biol Rev. 75: 519-631.
Jansen J, Friesema EC, Milici C, Visser TJ. (2005). Thyroid hormone transporters in health and disease. Thyroid. 15: 757-768.
Larsen PR. (2009).Type 2 iodothyronine deiodinase in human skeletal muscle: new insights into its physiological role and regulation. J Clin Endocrinol Metab. 94:1893-1895.
Liu XS, Cai Y, Wang Y, Xu SH, Ji K, Choi K. 2019. Effects of tris(1,3-dichloro-2-propyl) phosphate (tdcpp) and triphenyl phosphate (tpp) on sex-dependent alterations of thyroid hormones in adult zebrafish. Ecotoxicology and Environmental Safety. 170:25-32.
Liu YW, Chan WK. 2002. Thyroid hormones are important for embryonic to larval transitory phase in zebrafish. Differentiation. 70(1):36-45.
Manzon RG, Youson JH. (1997). The effects of exogenous thyroxine (T4) or triiodothyronine (T3), in the presence and absence of potassium perchlorate, on the incidence of metamorphosis and on serum T4 and T3 concentrations in larval sea lampreys (Petromyzon marinus L.). Gen Comp Endocrinol. 106:211-220.
Miller MD, Crofton KM, Rice DC, Zoeller RT. (2009).Thyroid-disrupting chemicals: interpreting upstream biomarkers of adverse outcomes. Environ Health Perspect. 117:1033-1041.
Moeller LC, Dumitrescu AM, Seo H, Refetoff S. (2006). Thyroid hormone mediated changes in gene expression can be initiated by cytosolic action of the thyroid hormone receptor β through the phosphatidylinositol 3-kinase pathway. NRS. 4:1-4.Nelson, K., Schroeder, A., Ankley, G., Blackwell, B., Blanksma, C., Degitz, S., Flynn, K., Jensen, K., Johnson, R., Kahl, M., Knapen, D., Kosian, P., Milsk, R., Randolph, E., Saari, T., Stinckens, E., Vergauwen, L., Villeneuve, D., 2016. Impaired anterior swim bladder inflation following exposure to the thyroid peroxidase inhibitor 2-mercaptobenzothiazole part I: Fathead minnow. Aquatic Toxicology 173, 192-203.Obregon MJ, Mallol J, Escobar del Rey F, Morreale de Escobar G. (1981). Presence of l-thyroxine and 3,5,3-triiodo-l-thyronine in tissues from thyroidectomised rats. Endocrinology 109:908-913.
Nelson K, Schroeder A, Ankley G, Blackwell B, Blanksma C, Degitz S, Flynn K, Jensen K, Johnson R, Kahl M et al. 2016. Impaired anterior swim bladder inflation following exposure to the thyroid peroxidase inhibitor 2-mercaptobenzothiazole part i: Fathead minnow. Aquatic Toxicology. 173:192-203.
Oetting A, Yen PM. (2007). New insights into thyroid hormone action. Best Pract Res Clin Endocrinol Metab. 21:193–208.
Power DM, Llewellyn L, Faustino M, Nowell MA, Bjornsson BT, Einarsdottir IE, Canario AV, Sweeney GE. 2001. Thyroid hormones in growth and development of fish. Comp Biochem Physiol C Toxicol Pharmacol. 130(4):447-459.
Ruuskanen S, Hsu BY. 2018. Maternal thyroid hormones: An unexplored mechanism underlying maternal effects in an ecological framework. Physiological and Biochemical Zoology. 91(3):904-916.
Santos CRA, Power DM. 1999. Identification of transthyretin in fish (sparus aurata): Cdna cloning and characterisation. Endocrinology. 140(5):2430-2433.
Schussler, G.C. (2000). The thyroxine-binding proteins. Thyroid 10:141–149.
Stinckens E, Vergauwen L, Blackwell BR, Anldey GT, Villeneuve DL, Knapen D. 2020. Effect of thyroperoxidase and deiodinase inhibition on anterior swim bladder inflation in the zebrafish. Environmental Science & Technology. 54(10):6213-6223.
Stinckens, E., Vergauwen, L., Schroeder, A., Maho, W., Blackwell, B., Witters, H., Blust, R., Ankley, G., Covaci, A., Villeneuve, D., Knapen, D., 2016. Impaired anterior swim bladder inflation following exposure to the thyroid peroxidase inhibitor 2-mercaptobenzothiazole part II: Zebrafish. Aquatic Toxicology 173, 204-217.
Thienpont B, Tingaud-Sequeira A, Prats E, Barata C, Babin PJ, Raldua D. 2011. Zebrafish eleutheroembryos provide a suitable vertebrate model for screening chemicals that impair thyroid hormone synthesis. Environmental Science & Technology. 45(17):7525-7532
Visser WE, Friesema EC, Jansen J, Visser TJ. (2008). Thyroid hormone transport in and out of cells. Trends Endocrinol Metab. 19:50-56.
Walpita CN, Van der Geyten S, Rurangwa E, Darras VM. 2007. The effect of 3,5,3'-triiodothyronine supplementation on zebrafish (danio rerio) embryonic development and expression of iodothyronine deiodinases and thyroid hormone receptors. Gen Comp Endocrinol. 152(2-3):206-214.
Wang JX, Shi GH, Yao JZ, Sheng N, Cui RN, Su ZB, Guo Y, Dai JY. 2020. Perfluoropolyether carboxylic acids (novel alternatives to pfoa) impair zebrafish posterior swim bladder development via thyroid hormone disruption. Environment International. 134.
Webster GM, Venners SA, Mattman A, Martin JW. 2014. Associations between perfluoroalkyl acids (pfass) and maternal thyroid hormones in early pregnancy: A population-based cohort study. Environmental Research. 133:338-347.
Williams GR. (2008). Neurodevelopmental and neurophysiological actions of thyroid hormone. J Neuroendocrinol. 20:784–794.
Yamauchi K, Nakajima J, Hayashi H, Hara A. 1999. Purification and characterization of thyroid-hormone-binding protein from masu salmon serum - a homolog of higher-vertebrate transthyretin. European Journal of Biochemistry. 265(3):944-949.
Yamauchi K1, Ishihara A. Evolutionary changes to transthyretin: developmentally regulated and tissue-specific gene expression.FEBS J. 2009 Oct;276(19):5357-66.
Yaoita Y, Brown DD. (1990). A correlation of thyroid hormone receptor gene expression with amphibian metamorphosis. Genes Dev. 4:1917-1924.
Yen PM. (2001). Physiological and molecular basis of thyroid hormone action. Physiol Rev. 81:1097-1142.
Zhang J, Grundstrom C, Brannstrom K, Iakovleva I, Lindberg M, Olofsson A, Andersson PL, Sauer-Eriksson AE. 2018. Interspecies variation between fish and human transthyretins in their binding of thyroid-disrupting chemicals. Environmental Science & Technology. 52(20):11865-11874.´
Zoeller RT, Tan SW, Tyl RW. General background on the hypothalamic-pituitary-thyroid (HPT) axis. Crit Rev Toxicol. 2007 Jan-Feb;37(1-2):11-53
Event: 1877: Altered, retinal layer structure
Short Name: Altered, retinal layer structure
Key Event Component
| Process | Object | Action |
|---|---|---|
| retina layer formation | retina | morphological change |
AOPs Including This Key Event
| AOP ID and Name | Event Type |
|---|---|
| Aop:363 - Thyroperoxidase inhibition leading to altered visual function via altered retinal layer structure | KeyEvent |
Stressors
| Name |
|---|
| Propylthiouracil |
| Methimazole |
| Perchlorate |
Biological Context
| Level of Biological Organization |
|---|
| Tissue |
Organ term
| Organ term |
|---|
| eye |
Evidence for Perturbation by Stressor
Propylthiouracil
6-n-propylthouracil is a classic positive control for inhibition of TPO.
Methimazole
Methimazole is a classic positive control for inhibition of TPO.
Perchlorate
Perchlorate ion (ClO− ₄) is a classic positive control for inhibition of NIS.
Domain of Applicability
Taxonomic Applicability| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| zebrafish | Danio rerio | High | NCBI |
| Life Stage | Evidence |
|---|---|
| Embryo | Moderate |
| Juvenile |
| Sex | Evidence |
|---|---|
| Unspecific | Moderate |
Taxonomic applicability: In general, the eye structure is very conserved among vertebrates, but some differences exist with regard to shape and expression of the different retinal layers. Fig. 1 (from Richardson 2012) demonstrates the histology of the human vs the zebrafish eye. As in humans, the mature zebrafish retina consists of three nuclear layers separated by two plexiform layers. The photoreceptor rod and cone nuclei are located in the outer nuclear layer; the amacrine, horizontal, and Müller glial cell bodies are found in the inner nuclear layer and the ganglion cell bodies are placed in the ganglion cell layer. The plexiform layers connect these layers. In contrast to zebrafish, the human retina lacks UV-sensitive cones.
Other structural differences between species are mostly related to their lifestyle (e.g. nocturnal vs diurnal) (Bibliowicz 2011) and cannot be generalized for specific vertebrate classes.
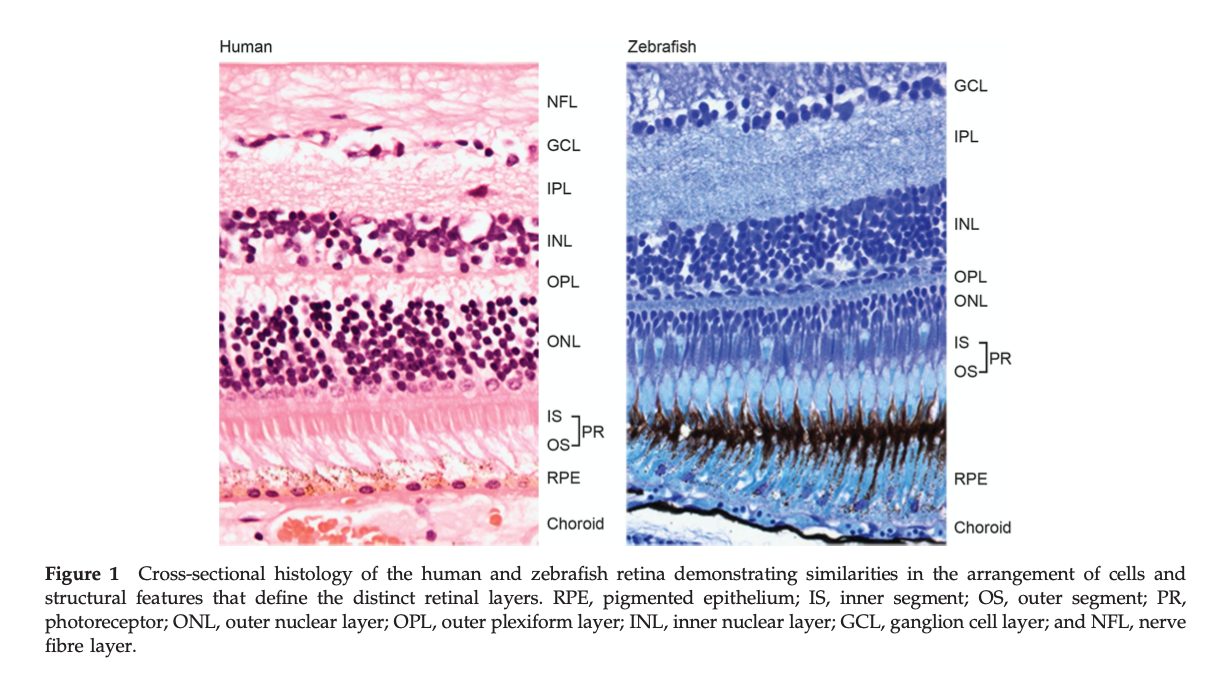
Life-stage applicability: Eye structure differs between life stages, as the different retinal layers do not develop at the same time and the eye itself grows with the organism. Eye development in zebrafish closely resembles the one in humans and other vertebrates. The eye develops from three different embryological tissues that form the specific structures of the eye, starting with the optic vesicle at 16 hpf, which further develops into the two-layered optic cup composed of the retinal neuroepithelium and pigmented epithelium until 20 hpf. Lens development begins as a lens placode that forms a solid lens mass by 22 hpf. Afterwards, the neuroectodermal layers of the optic vesicle invaginate ventrally by 24 hpf. By 48 hpf, zebrafish eye morphogenesis is almost complete with only retinal neurogenesis continuing. Retinal pigment epithelium flattening and final differentiation occurs around 27 hpf (Moreno-Marmol and others 2018). By 60 hpf, the different layers of the retina can be distinguished (Morris and Fadool 2005; Schmitt and Dowling 1999). Thereafter, further differentiation and maturation of the layers and cell types continues (Raymond and others 1995). For example, rods continue to mature until around 20 dpf (Morris and Fadool 2005). Impacts on retinal layer structure have been reported at 48, 66, 72, 96 and 120 hpf during zebrafish embryo-eleutheroembryo development (Baumann and others 2016; Komoike and others 2013; Reider and Connaughton 2014).
Sex applicability: There is no indication for sex differences in eye structure in fish or other vertebrates.
Evidence for perturbation by stressor: Multiple studies demonstrate that eye development and its resulting structure can be disrupted by different stressors (reviewed for example by Chen 2020).
Key Event Description
The anatomy and histology of the eye are highly conserved among vertebrates. The cornea and lens refract and focus light onto the posterior chamber of the eye, the vitreous cavity, which is covered by the retina. The retina consists of three specialised layers of cells, the outermost of which is formed by photoreceptors that absorb light and transmit the subsequent neural signal to the innermost layers, which consist of neurons specialised in processing and transmitting this neural signal (Wässle and Riemann, 1978; Cameron and Carney, 2000 ; Rockhill et al, 2000; Fadool, 2003). The neurons of the innermost layer converge to form the optic nerve, which transmits visual information to the brain (Gestri et al., 2012). The retina has different types of photoreceptors, the cones, which are responsible for colour vision, and the rods, which enable vision in the dark or in very low light conditions. In adults, cones are distributed in the retina in a precise and very regular arrangement, forming a photoreceptor mosaic. The precise spatiotemporal pattern of maturation of cones may affect the organization of this mosaic, and THs appear to play a role in the coordination of this maturation process (Suzuki et al., 2013). In the fish retina, this arrangement is most evident in the outer nuclear layer where the position of each cone subtype is precisely arranged relative to the others (Fadool, 2003; Robinson et al., 1993) resulting in a highly ordered crystalline-like mosaic.
The retinal pigment epithelium (RPE) is important to maintain a healthy and functional retina (Strauss 2005). The strong connection between the RPE cells with the tight junction, creates a blood-retinal barrier to mediate the directional transport of ions, water and nutrients while removing waste products. Another key function of the RPE is to absorb excess light energy to protect the neural retina from phototoxicity (Plafker 2012). Phagocytosis of spilled photoreceptor outer segments (Lister 2002) is another function of the RPE to maintain balanced photoreceptor growth, which is important for function.
Studies that detect and measure altered retinal layer structure after exposure to thyroid hormones or endocrine disruptors show, for example, altered cone cell number (Allison et al., 2006; Houbrechts et al., 2016; Vancamp et al., 2019), altered retinal cell number (Dong et al., 2014), or a general alteration of retinal morphology (Gamborino et al., 2001; Houbrechts et al., 2016; Komoike et al., 2013; Li et al., 2012; Reider & Connaughton, 2014), alteration of the pigment epithelium (Baumann et al., 2016), abnormal cone differentiation (Duval & Allison, 2018; Suzuki et al., 2013; Viets et al., 2016) or prevention of the opsin switch (Gan & Flamarique, 2010; Raine & Hawryshyn, 2009).
How it is Measured or Detected
For assessment of eye structure and layers, mostly simple morphometric analyses based on histological sections are sufficient. This can either be electron microscopy for subcellular changes, or normal light microscopy for cellular changes. Specific antibody staining might help to identify the different retinal layers, but usually, they are easily distinguishable by normal histological staining (e.g. HE staining).
Measurement of cell layer diameter is the most popular and simple method to assess changes in eye structure and layers. Moreover, measurement of the pigmentation grade of the retinal pigment epithelium can be used to assess structural changes.
(reviewed in Chen 2020)
References
Allison, W. T., Dann, S. G., Veldhoen, K. M., & Hawryshyn, C. W. (2006). Degeneration and regeneration of ultraviolet cone photoreceptors during development in rainbow trout. Journal of Comparative Neurology, 499(5), 702–715. https://doi.org/10.1002/cne.21164
Ali S, Champagne DL, Richardson MK. Behavioral profiling of zebrafish embryos exposed to a panel of 60 water-soluble compounds. Behav Brain Res. 2012;228(2):272-283. doi:10.1016/j.bbr.2011.11.020
Baumann, L., Ros, A., Rehberger, K., Neuhauss, S. C. F., & Segner, H. (2016). Thyroid disruption in zebrafish (Danio rerio) larvae: Different molecular response patterns lead to impaired eye development and visual functions.Aquatic Toxicology, 172, 44–55. https://doi.org/10.1016/j.aquatox.2015.12.015
Baumann, L., Segner, H., Ros, A., Knapen, D., & Vergauwen, L. (2019). Thyroid Hormone Disruptors Interfere with Molecular Pathways of Eye Development and Function in Zebrafish. International Journal of Molecular Sciences, 20(7), 1543. https://doi.org/10.3390/ijms20071543
Bibliowicz J, Tittle RK, Gross JM. Toward a Better Understanding of Human Eye Disease: Insights from the Zebrafish, Danio Rerio. Vol 100.; 2011. doi:10.1016/B978-0-12-384878-9.00007-8
Cameron, D.A., Carney, L.H., 2000. Cell mosaic patterns in the native and regenerated inner retina of zebrafish: implications for retinal assembly. J. Comp. Neurol. 416, 356–367.
Dong, W., Macaulay, L. J., Kwok, K. W., Hinton, D. E., Ferguson, P. L., & Stapleton, H. M. (2014). The PBDE metabolite 6-OH-BDE 47 affects melanin pigmentation and THRβMRNA expression in the eye of zebrafish embryos. Endocrine Disruptors, 2(1), e969072. https://doi.org/10.4161/23273739.2014.969072
Duval, M. G., & Allison, W. T. (2018). Photoreceptor progenitors depend upon coordination of gdf6a, thrβ, and tbx2b to generate precise populations of cone photoreceptor subtypes. Investigative Ophthalmology and Visual Science, 59(15), 6089–6101. https://doi.org/10.1167/iovs.18-24461
Fadool, J.M., 2003. Development of a rod photoreceptor mosaic revealed in transgenic zebrafish. Dev. Biol. 258, 277–290.
Gamborino, M. J., Sevilla-Romero, E., Muñoz, A., Hernández-Yago, J., Renau-Piqueras, J., & Pinazo-Durán, M. D. (2001). Role of thyroid hormone in craniofacial and eye development using a rat model. Ophthalmic Research,33(5), 283–291. https://doi.org/10.1159/000055682
Gan, K. J., & Flamarique, I. N. (2010). Thyroid hormone accelerates opsin expression during early photoreceptor differentiation and induces opsin switching in differentiated TRα-expressing cones of the salmonid retina. Developmental Dynamics, 239(10), 2700–2713. https://doi.org/10.1002/dvdy.22392
Gestri, G., Link, B. A., & Neuhauss, S. C. (2012). The visual system of zebrafish and its use to model
Houbrechts, A. M., Vergauwen, L., Bagci, E., Van houcke, J., Heijlen, M., Kulemeka, B., Hyde, D. R., Knapen, D., & Darras, V. M. (2016). Deiodinase knockdown affects zebrafish eye development at the level of gene expression, morphology and function. Molecular and Cellular Endocrinology, 424, 81–93. https://doi.org/10.1016/j.mce.2016.01.018
Komoike, Y., Matsuoka, M., & Kosaki, K. (2013). Potential teratogenicity of methimazole: Exposure of zebrafish embryos to methimazole causes similar developmental anomalies to human methimazole embryopathy. Birth Defects Research Part B - Developmental and Reproductive Toxicology, 98(3), 222–229. https://doi.org/10.1002/bdrb.21057
Li, Z., Ptak, D., Zhang, L., Walls, E. K., Zhong, W., & Leung, Y. F. (2012). Phenylthiourea specifically reduces zebrafish eye size. PLoS ONE,7(6), 1–14. https://doi.org/10.1371/journal.pone.0040132
Lister JA. Development of pigment cells in the zebrafish embryo. Microsc Res Tech. 2002;58(6):435-441. doi:10.1002/jemt.10161
Moreno-Marmol T, Cavodeassi F, Bovolenta P. 2018. Setting Eyes on the Retinal Pigment Epithelium. Frontiers in Cell and Developmental Biology 6.
Morris AC, Fadool JM. 2005. Studying rod photoreceptor development in zebrafish. Physiology & Behavior 86(3):306-313.
Plafker SM, O'Mealey GB, Szweda LI. Mechanisms for countering oxidative stress and damage in retinal pigment epithelium. Int Rev Cell Mol Biol. 2012;298:135-77. doi: 10.1016/B978-0-12-394309-5.00004-3. PMID: 22878106; PMCID: PMC3564215.
Raine, J. C., & Hawryshyn, C. W. (2009). Changes in thyroid hormone reception precede SWS1 opsin downregulation in trout retina. Journal of Experimental Biology, 212(17), 2781–2786. https://doi.org/10.1242/jeb.030866
Raymond PA, Barthel LK, Curran GA. 1995. DEVELOPMENTAL PATTERNING OF ROD AND CONE PHOTORECEPTORS IN EMBRYONIC ZEBRAFISH. Journal of Comparative Neurology 359(4):537-550.
Reider, M., & Connaughton, V. P. (2014). Effects of low-dose embryonic thyroid disruption and rearing temperature on the development of the eye and retina in zebrafish. Birth Defects Research. Part B, Developmental and Reproductive Toxicology, 101(5), 347–354. https://doi.org/10.1002/bdrb.21118
Rockhill, R.L., Euler, T., Masland, R.H., 2000. Spatial order within but not between types of retinal neurons. Proc. Natl. Acad. Sci. USA 97, 2303–2307.
Schmitt EA, Dowling JE. 1999. Early retinal development in the zebrafish, Danio rerio: Light and electron microscopic analyses. Journal of Comparative Neurology 404(4):515-536.
Suzuki, S. C., Bleckert, A., Williams, P. R., Takechi, M., Kawamura, S., & Wong, R. O. L. (2013). Cone photoreceptor types in zebrafish are generated by symmetric terminal divisions of dedicated precursors. Proceedings of the National Academy of Sciences, 110(37), 15109 LP – 15114.https://doi.org/10.1073/pnas.1303551110
Strauss O. The retinal pigment epithelium in visual function. Physiol Rev. 2005 Jul;85(3):845-81. doi: 10.1152/physrev.00021.2004. PMID: 15987797.
Vancamp, P., Bourgeois, N. M. A., Houbrechts, A. M., & Darras, V. M. (2019). Knockdown of the thyroid hormone transporter MCT8 in chicken retinal precursor cells hampers early retinal development and results in a shift towards more UV/blue cones at the expense of green/red cones. Experimental Eye Research, 178(September 2018), 135–147. https://doi.org/10.1016/j.exer.2018.09.018
Viets, K., Eldred, K. C., & Johnston, R. J. (2016). Mechanisms of Photoreceptor Patterning in Vertebrates and Invertebrates. Trends in Genetics, 32(10), 638–659. https://doi.org/10.1016/j.tig.2016.07.004
Wässle, H., Riemann, H.J., 1978. The mosaic of nerve cells in the mammalian retina. Proc. R. Soc. London B Biol. Sci. 200, 441–461.
List of Adverse Outcomes in this AOP
Event: 1643: Altered, Visual function
Short Name: Altered, Visual function
Key Event Component
| Process | Object | Action |
|---|---|---|
| vision trait | eye | functional change |
AOPs Including This Key Event
Stressors
| Name |
|---|
| Propylthiouracil |
Biological Context
| Level of Biological Organization |
|---|
| Organ |
Organ term
| Organ term |
|---|
| eye |
Domain of Applicability
Taxonomic Applicability| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| zebrafish | Danio rerio | NCBI |
| Life Stage | Evidence |
|---|---|
| Embryo | High |
| Juvenile | Moderate |
| Sex | Evidence |
|---|---|
| Unspecific |
-
Taxonomic applicability
Visual function decrease can be evaluated in wide range of species including mammals, amphibians, fish and humans. Evaluation of these visual function modification change according to the species and its environment.
-
Life-stage applicability
Vision plays a crucial role in the early life stages of most species, as timing of eye development and establishment of functional vision is essential for perception of food or avoidance of predators for example (Carvalho et al., 2002).The first visual responses based on retinal functionality appear around 70 hpf in zebrafish (Schmitt and Dowling 1999). It is plausible to assume that alterations of the eye structure would result in altered visual function across all life stages, but such alterations are most likely to occur during the development of the normal eye structure, which occurs in the embryo-eleutheroembryo phase. Some studies have also shown a decrease in vision related to age (Brastrom et al., 2019; Martínez-Roda et al., 2016; Segura et al., 2018) including on visual acuity, visual fields, colour vision and dark adaptation, are well documented (Hennelly et al, 1998).
-
Sex applicability
Sex seems not relevant for most of the visual function decreases observed in different studies. Differences according to the sex of the individuals have however been reported concerning the basic visual capacities (e.g. color perception, contrast sensitivity, visual acuity, motion perception,...) but also concerning the frequency of certain diseases influencing these diminished visual functions, notably in humans (Vanston and Strother, 2017).
Key Event Description
The decrease in visual function can have different aspects, such as loss of chromatic vision, changes in eye movements, differences in sensitivity to light, but also changes in the retinal pigment epithelium (RPE) that may be related to a decrease in visual function (Strauss, 2005). The visual system is highly variable from one species to another, and this variability is a key factor influencing animal behaviour (Corral-López et al., 2017).
Decreases in these visual functions can have a strong impact on behaviour, leading to changes in individual response and abilities in the environment, including, for example, perception of food or avoidance of predators. Variation in the visual system can also influence learning tasks when visual stimuli are used (Corral-López et al., 2017).
Some studies detect visual impairments, such as an optokinetic response decrease and increase of light preference of zebrafish larvae after treatment with the endocrine disruptor propylthiouracil (Baumann et al 2016 ), after chronic dietary selenomethionine exposure (Raine et al 2016), exposure to PCBs (Zhang et al, 2015) or deiodinase knockdown (Houbrechts et al 2016, Vancamp et al 2018).
How it is Measured or Detected
Studies of spatial and temporal aspects of the visual system have focused on assessments of optomotor (whole body swimming) or optokinetic (eye movement) responses. These responses are unconditioned optical and motor reactions including:
-
Opto Kinetic response, OKR (similar protocol for Rat/mice (Segura et al., 2018), fish (Zou et al., 2010) and humans (Kang and Wildsoet, 2016)). The OKR is a visually-mediated assay in which an individual will respond to alternating black and white stripes by exhibiting eye saccades, eye movements without coordinated body movements, in the same direction as rotating stripes. An eye saccade relies on the ability to rapidly move the eye from focusing on one external target to the next in a repeated manner (Magnuson et al., 2020). Optokinetic tracking has a robust performance and does not require training the animal, allowing for the quick assessment (and at earlier ages) of visual features such as visual acuity (VA) and contrast sensitivity (CS)11–14. However, the main disadvantage of optokinetic tracking is that it is a subjective method in which the decision about whether the animal is performing the optokinetic tracking or not is made by an experimenter (Segura et al., 2018).
-
Opto Motor Reponses, OMR. OMR tracks the ability of the fish to swim in the direction of a perceived motion when presented with a whole-field stimulus (Neuhauss, 2003), (Gould et al., 2017)).
-
Light-dark transition or vision startle response: reaction to change in light intensity (light sensitivity) (Brastrom et al., 2019)
-
Black-white preference test (Baumann et al., 2016)
-
Diverse Mobility assay including Tracking, touch-evoked escape-response assays, Swirl assays, locomotion assay, swimming activity, phototactic swimming activity assay, induced locomotor response (LLR) (Baumann et al., 2016; Gao et al., 2015; Zhao et al., 2014))
References
Baumann, L., Ros, A., Rehberger, K., Neuhauss, S. C. F., & Segner, H. (2016). Thyroid disruption in zebrafish (Danio rerio) larvae: Different molecular response patterns lead to impaired eye development and visual functions. Aquatic Toxicology, 172, 44–55. https://doi.org/10.1016/j.aquatox.2015.12.015
Brastrom, L.K., Scott, C.A., Dawson, D. V., Slusarski, D.C., 2019. A High-Throughput Assay for Congenital and Age-Related Eye Diseases in Zebrafish. Biomedicines 7, 28. https://doi.org/10.3390/biomedicines7020028
Carvalho, P.S.M., Noltie, D.B., Tillitt, D.E., 2002. Ontogenetic improvement of visual function in the medaka Oryzias latipes based on an optomotor testing system for larval and adult fish. Anim. Behav. 64, 1–10. https://doi.org/10.1006/anbe.2002.3028
Corral-López, A., Garate-Olaizola, M., Buechel, S.D., Kolm, N., Kotrschal, A., 2017. On the role of body size, brain size, and eye size in visual acuity. Behav. Ecol. Sociobiol. 71. https://doi.org/10.1007/s00265-017-2408-z
Gao, D., Wu, M., Wang, C., Wang, Y., Zuo, Z., 2015. Chronic exposure to low benzo[a]pyrene level causes neurodegenerative disease-like syndromes in zebrafish (Danio rerio). Aquat. Toxicol.
Houbrechts, A. M., Vergauwen, L., Bagci, E., Van houcke, J., Heijlen, M., Kulemeka, B., Hyde, D. R., Knapen, D., & Darras, V. M. (2016). Deiodinase knockdown affects zebrafish eye development at the level of gene expression, morphology and function. Molecular and Cellular Endocrinology, 424, 81–93. https://doi.org/10.1016/j.mce.2016.01.018
Raine, J. C., Lallemand, L., Pettem, C. M., & Janz, D. M. (2016). Effects of Chronic Dietary Selenomethionine Exposure on the Visual System of Adult and F1 Generation Zebrafish (Danio rerio). Bulletin of Environmental Contamination and Toxicology, 97(3), 331–336. https://doi.org/10.1007/s00128-016-1849-9
Vancamp, P., Bourgeois, N. M. A., Houbrechts, A. M., & Darras, V. M. (2019). Knockdown of the thyroid hormone transporter MCT8 in chicken retinal precursor cells hampers early retinal development and results in a shift towards more UV/blue cones at the expense of green/red cones. Experimental Eye Research,178(September 2018), 135–147. https://doi.org/10.1016/j.exer.2018.09.018
Zhang, X., Hong, Q., Yang, L., Zhang, M., Guo, X., Chi, X., & Tong, M. (2015). PCB1254 exposure contributes to the abnormalities of optomotor responses and influence of the photoreceptor cell development in zebrafish larvae. Ecotoxicology and Environmental Safety, 118, 133–138. https://doi.org/10.1016/j.ecoenv.2015.04.026
Event: 351: Increased Mortality
Short Name: Increased Mortality
Key Event Component
| Process | Object | Action |
|---|---|---|
| mortality | increased |
AOPs Including This Key Event
Biological Context
| Level of Biological Organization |
|---|
| Population |
Domain of Applicability
Taxonomic Applicability| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| all species | all species | High | NCBI |
| Life Stage | Evidence |
|---|---|
| All life stages | High |
| Sex | Evidence |
|---|---|
| Unspecific | Moderate |
All living things are susceptible to mortality.
Key Event Description
Increased mortality refers to an increase in the number of individuals dying in an experimental replicate group or in a population over a specific period of time.
How it is Measured or Detected
Mortality of animals is generally observed as cessation of the heart beat, breathing (gill or lung movement) and locomotory movements. Mortality is typically measured by observation. Depending on the size of the organism, instruments such as microscopes may be used. The reported metric is mostly the mortality rate: the number of deaths in a given area or period, or from a particular cause.
Depending on the species and the study setup, mortality can be measured:
- in the lab by recording mortality during exposure experiments
- in dedicated setups simulating a realistic situation such as mesocosms or drainable ponds for aquatic species
- in the field, for example by determining age structure after one capture, or by capture-mark-recapture efforts. The latter is a method commonly used in ecology to estimate an animal population's size where it is impractical to count every individual.
Regulatory Significance of the AO
Increased mortality is one of the most common regulatory assessment endpoints, along with reduced growth and reduced reproduction.
Event: 360: Decrease, Population trajectory
Short Name: Decrease, Population trajectory
Key Event Component
| Process | Object | Action |
|---|---|---|
| population growth rate | population of organisms | decreased |
AOPs Including This Key Event
Biological Context
| Level of Biological Organization |
|---|
| Population |
Domain of Applicability
Taxonomic Applicability| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| all species | all species | High | NCBI |
| Life Stage | Evidence |
|---|---|
| All life stages | Not Specified |
| Sex | Evidence |
|---|---|
| Unspecific | Not Specified |
Consideration of population size and changes in population size over time is potentially relevant to all living organisms.
Key Event Description
Population ecology is the study of the sizes (and to some extent also the distribution) of plant and animal populations and of the processes, mainly biological in nature, that determine these sizes. As such, it provides an integrated measure of events occurring at lower levels of biological organization (biochemical, organismal, etc.). The population size in turn determines community and ecosystem structure. For fish, maintenance of sustainable fish and wildlife populations (i.e., adequate to ensure long-term delivery of valued ecosystem services) is an accepted regulatory goal upon which risk assessments and risk management decisions are based.
How it is Measured or Detected
Population trajectories, either hypothetical or site specific, can be estimated via population modeling based on measurements of vital rates or reasonable surrogates measured in laboratory studies. As an example, Miller and Ankley 2004 used measures of cumulative fecundity from laboratory studies with repeat spawning fish species to predict population-level consequences of continuous exposure.
Regulatory Significance of the AO
Maintenance of sustainable fish and wildlife populations (i.e., adequate to ensure long-term delivery of valued ecosystem services) is a widely accepted regulatory goal upon which risk assessments and risk management decisions are based.
References
- Miller DH, Ankley GT. 2004. Modeling impacts on populations: fathead minnow (Pimephales promelas) exposure to the endocrine disruptor 17ß-trenbolone as a case study. Ecotoxicology and Environmental Safety 59: 1-9.
Appendix 2
List of Key Event Relationships in the AOP
List of Adjacent Key Event Relationships
Relationship: 309: Thyroperoxidase, Inhibition leads to TH synthesis, Decreased
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Inhibition of Thyroperoxidase and Subsequent Adverse Neurodevelopmental Outcomes in Mammals | adjacent | High | Low |
| Thyroperoxidase inhibition leading to increased mortality via reduced anterior swim bladder inflation | adjacent | High | Low |
| Inhibition of thyroid peroxidase leading to impaired fertility in fish | adjacent | High | High |
| Thyroperoxidase inhibition leading to altered amphibian metamorphosis | adjacent | High | Moderate |
| Thyroperoxidase inhibition leading to altered visual function via altered retinal layer structure | adjacent | High | Low |
| Thyroperoxidase inhibition leading to altered visual function via decreased eye size | adjacent | ||
| Thyroperoxidase inhibition leading to altered visual function via altered photoreceptor patterning | adjacent | ||
| Inhibition of thyroid peroxidase leading to follicular cell adenomas and carcinomas (in rat and mouse) | adjacent |
Evidence Supporting Applicability of this Relationship
| Life Stage | Evidence |
|---|---|
| All life stages | High |
| Sex | Evidence |
|---|---|
| Male | High |
| Female | High |
Taxonomic: This KER is plausibly applicable across vertebrates. Inhibition of TPO activity is widely accepted to directly impact TH synthesis. This is true for both rats and humans, as well as some fishes, frogs and birds. Most of the data supporting a causative relationship between TPO inhibition and altered TH synthesis is derived from animal studies, in vitro thyroid microsomes from rats or pigs, and a limited number of human ex vivo (Nagasaka and Hidaka, 1976; Vickers et al., 2012) and clinical studies. There are data to support that gene mutations in TPO result in congenital hypothyroidism, underscoring the essential role of TPO in human thyroid hormone synthesis.
Life stage: Applicability to certain life stages may depend on the species and their dependence on maternally transferred thyroid hormones during the earliest phases of development. The earliest life stages of teleost fish rely on maternally transferred THs to regulate certain developmental processes until embryonic TH synthesis is active (Power et al., 2001). As a result, TPO inhibition is not expected to decrease TH synthesis during these earliest stages of development. In zebrafish, Opitz et al. (2011) showed the formation of a first thyroid follicle at 55 hours post fertilization (hpf), Chang et al. (2012) showed a first significant TH increase at 120 hpf and Walter et al. (2019) showed clear TH production already at 72 hpf but did not analyse time points between 24 and 72 hpf. In fathead minnows, a significant increase of whole body thyroid hormone levels was already observed between 1 and 2 dpf, which corresponds to the appearance of the thyroid anlage at 35 hpf prior to the first observation of thyroid follicles at 58 hpf (Wabuke-Bunoti and Firling, 1983). It is still uncertain when exactly embryonic TH synthesis is activated and how this determines sensitivity to TH disruptors.
Sex: The KE is plausibly applicable to both sexes. Thyroid hormones are essential in both sexes and the components of the HPT-axis are identical in both sexes. There can however be sex-dependent differences in the sensitivity to the disruption of thyroid hormone levels and the magnitude of the response. In humans, females appear more susceptible to hypothyroidism compared to males when exposed to certain halogenated chemicals (Hernandez‐Mariano et al., 2017; Webster et al., 2014). In adult zebrafish, Liu et al. (2019) showed sex-dependent changes in thyroid hormone levels and mRNA expression of regulatory genes including corticotropin releasing hormone (crh), thyroid stimulating hormone (tsh) and deiodinase 2 after exposure to organophosphate flame retardants. The underlying mechanism of any sex-related differences remains unclear.
Key Event Relationship Description
Thyroperoxidase (TPO) is a heme-containing apical membrane protein within the follicular lumen of thyrocytes that acts as the enzymatic catalyst for thyroid hormone (TH) synthesis (Taurog, 2005) across vertebrates. Two commonly used reference chemicals, propylthiouracil (PTU) and methimazole (MMI), are drugs that inhibit the ability of TPO to: a) activate iodine and transfer it to thyroglobulin (Tg) (Davidson et al., 1978); and, b) couple thyroglobulin (Tg)-bound iodotyrosyls to produce Tg-bound thyroxine (T4) and triiodothyronine (T3) (Taurog, 2005).
Evidence Supporting this KER
The weight of evidence supporting a direct linkage between the MIE, TPO inhibition, and the KE of decreased TH synthesis, is strong and supported by more than three decades of research in animals, including humans (Cooper et al., 1982; Cooper et al.,1983; Divi and Doerge, 1994).
Biological PlausibilityThe biological plausibility for this KER is rated Strong. TPO is the only enzyme capable of de novo systhesis of TH. TPO catalyzes several reactions, including the oxidation of iodide, nonspecific iodination of tyrosyl residues of thyroglobulin (Tg) to form monoiodotyrosyl (MIT) or diiodotyrosyl (DIT) residues, and the coupling of these Tg-bound iodotyrosyls to produce Tg-bound T3 and T4 (Divi and Doerge, 1994; Kessler et al., 2008; Ruf et al., 2006; Taurog et al., 1996, 2005). Therefore, inhibition of TPO activity is widely accepted to directly impact TH synthesis.
Empirical EvidenceEmpirical support for this KER is strong. There are several papers that have measured alterations in TPO and subsequent effects on TH synthesis across vertebrates. Taurog et al. (1996) showed decreased guicaol activity, decreased bound I125, and subsequent decreases in newly formed T3 and T4 per molecule of Tg, following exposure to PTU, MMI and some antibiotics. There is important evidence in mammals. Following in vivo exposure to PTU in rats (Cooper et al., 1982; 1983), there are concentration and time-dependent decreases in thyroid protein bound iodine and serum T4 and T3 that recovered one month after cessation of PTU exposure. In addition, measures of thyroidal iodine content were highly correlated with intra-thyroidal PTU concentration. Vickers et al. (2012) demonstrated dose- and time- dependent inhibition of TPO activity in both human and rat thyroid homogenates exposed to MMI.
Tietge et al (2010) showed decreases in thyroidal T4 following MMI exposure in Xenopus. Also in Xenopus, Haselman et al (2020) showed decreases in thyroidal iodotyrosines (MIT/DIT) and iodothyronines (T4/T3) following exposure to MMI. Doerge et al (1998) showed that a tryphenylmethane dye, malachite green, inhibited TPO and lowered thyroxine production. A recent paper used a series of benzothiazoles and showed TPO inhibition (guicaol assay) and inhibition of TSH stimulated thyroxine release from Xenopus thyroid gland explant cultures (Hornung et al., 2015).
Additionally, evidence is available from studies investigating responses to TPO inhibitors in fish. For example, Stinckens et al. (2020) showed reduced whole body T4 concentrations in zebrafish larvae exposed to 50 or 100 mg/L methimazole, a potent TPO inhibitor, from immediately after fertilization until 21 or 32 days of age. Exposure to 37 or 111 mg/L propylthiouracil also reduced T4 levels after exposure up to 14, 21 and 32 days in the same study. Walter et al. (2019) showed that propylthiouracil had no effect on T4 levels in 24h old zebrafish, but decreased T4 levels of 72h old zebrafish. This difference is probably due to the onset of embryonic TH production between the age of 24 and 72 hours (Opitz et al., 2011). Stinckens et al. (2016) showed that exposure to 2-mercaptobenzothiazole (MBT), an environmentally relevant TPO inhibitor, decreased whole body T4 levels in continuously exposed 5 and 32 day old zebrafish larvae. Several other studies have also shown that chemically induced Inhibition of TPO results in reduced TH synthesis in zebrafish (Van der Ven et al., 2006; Raldua and Babin, 2009; Liu et al., 2011; Thienpont et al., 2011; Rehberger et al., 2018). A high concentration of MBT also decreased whole body T4 levels in 6 day old fathead minnows, but recovery was observed at the age of 21 days although the fish were kept in the exposure medium (Nelson et al., 2016). Crane et al. (2006) showed decreased T4 levels in 28 day old fathead minnows continuously exposed to 32 or 100 µg/L methimazole.
Temporal Evidence: In mammals, the temporal nature of this KER is applicable to all life stages, including development (Seed et al., 2005). The impact of decreased TPO activity on thyroidal hormone synthesis is similar across all ages in mammals. Good evidence for the temporal relationship of the KER comes from thyroid system modeling (e.g., Degon et al., 2008; Fisher et al., 2013) using data from studies of iodine deficiency and chemicals that inhibit NIS. In addition, there is ample evidence of the temporal impacts of TPO inhibition on TH synthesis, using ex vivo and in vitro measures that demonstrate the time course of inhibition following chemical exposures, including some data from human thyroid microsomes and ex vivo thyroid slices (Vickers et al., 2012). Future work is needed that measures both TPO inhibition and TH production during development.
In oviparous fish such as zebrafish and fathead minnow, the nature of this KER depends on the life stage since the earliest stages of embryonic development rely on maternal thyroid hormones transferred to the eggs. Embryonic thyroid hormone synthesis is activated later during embryo-larval development. (See Domain of applicability)
Dose-Response Evidence: Dose-response data is available from a number of studies in mammals that correlate TPO inhibition with decreased TH production measured using a variety of endpoints including iodine organification (e.g., Taurog et al., 1996), inhibition of guicaol oxidation in thyroid microsomes (e.g., Doerge and Chang, 2002), and direct measure of thyroid gland T4 concentrations (e.g., Hornung et al., 2015). However, there is a lack of dose-response data from developmental studies showing direct linkages from TPO inhibition to thyroidal TH synthesis.
Uncertainties and InconsistenciesWhile it is clear that TPO inhibition will lead to altered hormone synthesis, there is a need for data that will inform quantitative modeling of the relationship between TPO inhibition and the magnitude of effects on thyroid hormone synthesis.
Data from studies on genistein highlight this uncertainty. Doerge and colleagues have demonstrated that for this compound up to 80% TPO inhibition did not result in decreased serum T4 in rats (Doerge and Chang, 2002). This is not consistent with other prototypical TPO inhibitors (e.g., PTU, MMI). Genistein is however a well-known phytoestrogen and the observed inconsistency may be the result of feedback mechanisms resulting from its estrogenic effect.
Quantitative Understanding of the Linkage
In Xenopus laevis, Haselman et al.(2020) demonstrated temporal profiles of thyroidal iodotyrosines (MIT/DIT) and iodothyronines (T4/T3), the products of TPO activity, following exposure to three different model TPO inhibitors (MMI, PTU, MBT) at multiple concentrations. This study established that, in Xenopus, measurable decreases in the products of TPO activity can occur as early as 2 days of exposure during pro-metamorphosis. However, despite consistent profiles of some iodo-species across chemicals, other iodo-species showed inconsistent profiles across chemicals. This highlights the multiple mechanisms of TPO (iodination and coupling) and differential susceptiblity to inhibition of those mechanisms depending on the chemical's type of interaction with TPO. The most consistent concentration-response relationship across chemicals and over time was demonstrated by thyroidal T4, which is the most relevant product to subsequent key events. At the highest concentrations tested for each chemcial, thyroidal T4 was below detection by 7 days of exposure across all three TPO inhibitors. Keeping in mind that the thyroid gland has follicular lumen space where thyroglobulin/T4 is stored until proteolysis and release to the blood, full inhibition of TPO would result in a delayed measureable response due to the time it takes to deplete stored hormone. Regardless of the delay, the results from this study imply full inhibition of TPO by each of these three chemicals at the highest test concentrations, but would require chemical residue analysis and/or toxicokinetic modeling to relate cellular/tissue concentrations at the site of TPO catalysis to levels of inhibition via Michaelis-Menten kinetic descriptions.
Profiles of thyroidal iodinated species demonstrated by Haselman et al. (2020) across three different TPO inhibitors suggests that a high level of TPO inhibition must occur in order to elicit responses in subsequent key events. Although the level of TPO inhibition is not directly quantifiable from this study, these data suggest that at least 90-100% inhibition was occurring since circulating T4 was not detectable at 10 days of exposure to the highest concentrations of MMI and MBT. However, additional efforts would be necessary to determine the minimum level of TPO inhibition that leads to a measurable decrease in thyroidal T4 and subsequently circulating T4.
Response-response relationshipThere are only a limited number of studies where both TPO inhibition and iodine organification have been measured in vivo, and there are not enough data available to make any definitive quantitative correlations. One in vivo study in rats exposed to the TPO inhibitor genistein found no in vivo impact on serum thyroid hormone concentrations, even when TPO was inhibited up to 80% (Chang and Doerge, 2000). Genistein is however a well-known phytoestrogen and the observed inconsistency may be the result of feedback mechanisms resulting from its estrogenic effect.
Given that this is an MIE to KE relationship, there is only one response to evaluate in the relationship. Decreased TH synthesis, as measured by responses of iodinated species in the thyroid gland, is the result of TPO inhibition, which cannot be measured directly in vivo.
Time-scaleIn vivo, evaluations of TPO inhibition are limited to evaluation of the iodinated species, or products of TPO activity, present in the thyroid gland at a particular time. However, as stated previously, any measurable response in these iodinated species is not a discreet assessment of TPO activity given that the gland maintains storage of hormone in the follicular lumen space and any alteration of TPO activity would be detected once the stores begin to be depleted. In Xenopus laevis, Haselman et al. (2020) showed a decrease in thyroidal iodinated species after only 2 days of exposure to potent TPO inhibitor MMI during thyroid-mediated metamorphosis and within 4 days for PTU and MBT, both model TPO inhibitors. In zebrafish, Walter et al. (2019) reported a similar time frame, namely a decrease in T4 levels at 72 hpf after starting the exposure to PTU at 0-2 hpf. It should be noted that the time-scale is probably depending on the developmental stage and whether the embryo is capable of thyroid hormone synthesis, rather than on the exposure duration.
Known modulating factorsIodine availability will impact the ability of TPO to iodinate tyrosine residues on thyroglobulin. Iodine availability to TPO can be impacted a number of ways. First, environmental availability of iodine can vary greatly depending on whether and how much iodine exists in surface waters for aquatic organisms (gill respirators) and in the diets of both terrestrial and aquatic organisms. Second, somewhat regardless of iodine availability through environmental uptake (i.e., barring extremely high iodine exposure), iodine is actively transported into the thyroid follicular cell from the blood via sodium-iodide symporter (NIS), which has been shown to be susceptible to inhibition by, for example, perchlorate. As such, iodine availability to TPO is mediated by functional NIS. Finally, iodine is not fully available to TPO on the apical surface of the thyroid follicular cell until it is transported through the apical membrane by pendrin, an anion exchange protein - mutations or inhibition of pendrin could affect iodine availability to TPO.
Hydrogen peroxide is also needed by TPO to mediate the oxidation of iodide, which is produced locally by dual oxidase (DUOX). A mutation or inhibition of DUOX will impact local production of H2O2 leading to lower oxidizing potential of TPO and less organification of iodide.
Known Feedforward/Feedback loops influencing this KERThyroid stimulating hormone (TSH) released from the pituitary positively regulates the synthesis and release of thyroid hormones from the thyroid gland. As such, when TPO is inhibited and thyroid hormone synthesis is decreased, lower systemic levels of hormone cause feedback from the pituitary via TSH to upregulate a number of processes in the thyroid gland as a means of compensation, including (but not limited to) enhanced gene expression of NIS and thyrocyte cell proliferation (Tietge et al., 2010; Haselman et al., 2020).
References
Chang HC, Doerge DR. Dietary genistein inactivates rat thyroid peroxidase in vivo without an apparent hypothyroid effect. Toxicol Appl Pharmacol 168:244–252 (2000).
Chang, J., Wang, M., Gui, W., Zhao, Y., Yu, L., Zhu, G., 2012. Changes in Thyroid Hormone Levels during Zebrafish Development. Zoological Science 29, 181-184.
Cooper DS, Kieffer JD, Halpern R, Saxe V, Mover H, Maloof F, Ridgway EC (1983) Propylthiouracil (PTU) pharmacology in the rat. II. Effects of PTU on thyroid function. Endocrinology 113:921-928.
Cooper DS, Saxe VC, Meskell M, Maloof F, Ridgway EC. Acute effects of propylthiouracil (PTU) on thyroidal iodide organification and peripheral iodothyronine deiodination: correlation with serum PTU levels measured by radioimmunoassay. J Clin Endocrinol Metab. 1982 54(1):101-7.
Crane HM, Pickford DB, Hutchinson TH, Brown JA. 2006. The effects of methimazole on development of the fathead minnow, pimephales promelas, from embryo to adult. Toxicological Sciences. 93(2):278-285.
Davidson, B., Soodak, M., Neary, J.T., Strout, H.V., and Kieffer, J.D. (1978). The irreversible inactivation of thyroid peroxidase by methylmercaptoimidazole, thiouracil, and propylthiouracil in vitro and its relationship to in vivo findings. Endocrinology 103:871–882.
Divi, R. L., and Doerge, D. R. (1994). Mechanism-based inactivation of lactoperoxidase and thyroid peroxidase by resorcinol derivatives. Biochemistry 33(32), 9668-74.
Doerge DR, Chang HC, Divi RL, Churchwell Mechanism for inhibition of thyroid peroxidase by leucomalachite green. Chem Res Toxicol. 1998 11(9):1098-104.
Doerge DR, Chang HC. Inactivation of thyroid peroxidase by soy isoflavones, in vitro and in vivo. J Chromatogr B Analyt Technol Biomed Life Sci. 2002 Sep 25;777(1-2):269-79
Haselman, J.T., Olker, J.H., Kosian, P.A., Korte, J.J., Swintek, J.A., Denny, J.S., Nichols, J.W., Tietge, J.E., Hornung, M.W. and Degitz, S.J., 2020. Targeted pathway-based in vivo testing using thyroperoxidase inhibition to evaluate plasma thyroxine as a surrogate metric of metamorphic success in model amphibian Xenopus laevis. Toxicological Sciences, 175(2), pp.236-250.
Hernandez-Mariano JA, Torres-Sanchez L, Bassol-Mayagoitia S, Escamilla-Nunez M, Cebrian ME, Villeda-Gutierrez EA, Lopez-Rodriguez G, Felix-Arellano EE, Blanco-Munoz J. 2017. Effect of exposure to p,p '-dde during the first half of pregnancy in the maternal thyroid profile of female residents in a mexican floriculture area. Environmental Research. 156:597-604.
Hornung MW, Kosian PA, Haselman JT, Korte JJ, Challis K, Macherla C, Nevalainen E, Degitz SJ. In Vitro, Ex Vivo, and In Vivo Determination of Thyroid Hormone Modulating Activity of Benzothiazoles.Toxicol Sci. 2015 146(2):254-64.
Kessler, J., Obinger, C., and Eales, G. (2008). Factors influencing the study of peroxidase-generated iodine species and implications for thyroglobulin synthesis. Thyroid 18(7), 769-74, 10.1089/thy.2007.0310.
Liu CS, Zhang XW, Deng J, Hecker M, Al-Khedhairy A, Giesy JP, Zhou BS. 2011. Effects of prochloraz or propylthiouracil on the cross-talk between the hpg, hpa, and hpt axes in zebrafish. Environmental Science & Technology. 45(2):769-775.
Liu XS, Cai Y, Wang Y, Xu SH, Ji K, Choi K. 2019. Effects of tris(1,3-dichloro-2-propyl) phosphate (tdcpp) and triphenyl phosphate (tpp) on sex-dependent alterations of thyroid hormones in adult zebrafish. Ecotoxicology and Environmental Safety. 170:25-32.
Nagasaka, A., and Hidaka, H. (1976). Effect of antithyroid agents 6-propyl-2-thiouracil and 1-mehtyl-2-mercaptoimidazole on human thyroid iodine peroxidase. J. Clin. Endocrinol. Metab. 43:152–158.
Nelson K, Schroeder A, Ankley G, Blackwell B, Blanksma C, Degitz S, Flynn K, Jensen K, Johnson R, Kahl M et al. 2016. Impaired anterior swim bladder inflation following exposure to the thyroid peroxidase inhibitor 2-mercaptobenzothiazole part i: Fathead minnow. Aquatic Toxicology. 173:192-203.
Opitz, R., Maquet, E., Zoenen, M., Dadhich, R., Costagliola, S., 2011. TSH Receptor Function Is Required for Normal Thyroid Differentiation in Zebrafish. Molecular Endocrinology 25, 1579-1599.
Power, D.M., Llewellyn, L., Faustino, M., Nowell, M.A., Bjornsson, B.T., Einarsdottir, I.E., Canario, A.V., Sweeney, G.E., 2001. Thyroid hormones in growth and development of fish. Comp Biochem Physiol C Toxicol Pharmacol 130, 447-459.
Raldua, D., Babin, P.J., 2009. Simple, Rapid Zebrafish Larva Bioassay for Assessing the Potential of Chemical Pollutants and Drugs to Disrupt Thyroid Gland Function. Environmental Science & Technology 43, 6844-6850.
Rehberger, K., Baumann, L., Hecker, M., Braunbeck, T., 2018. Intrafollicular thyroid hormone staining in whole-mount zebrafish (Danio rerio) embryos for the detection of thyroid hormone synthesis disruption. Fish Physiology and Biochemistry 44, 997-1010.
Ruf, J., and Carayon, P. (2006). Structural and functional aspects of thyroid peroxidase. Archives of biochemistry and biophysics 445(2), 269-77, 10.1016/j.abb.2005.06.023.
Stinckens E, Vergauwen L, Blackwell BR, Anldey GT, Villeneuve DL, Knapen D. 2020. Effect of thyroperoxidase and deiodinase inhibition on anterior swim bladder inflation in the zebrafish. Environmental Science & Technology. 54(10):6213-6223.
Stinckens E, Vergauwen L, Schroeder A, Maho W, Blackwell B, Witters H, Blust R, Ankley G, Covaci A, Villeneuve D et al. 2016. Impaired anterior swim bladder inflation following exposure to the thyroid peroxidase inhibitor 2-mercaptobenzothiazole part ii: Zebrafish. Aquatic Toxicology. 173:204-217.
Taurog A. 2005. Hormone synthesis. In: Werner and Ingbar’s The Thyroid: A Fundamental and Clinical Text (Braverman LE, Utiger RD, eds). Philadelphia:Lippincott, Williams and Wilkins, 47–81.
Taurog, A., Dorris, M. L., and Doerge, D. R. (1996). Mechanism of simultaneous iodination and coupling catalyzed by thyroid peroxidase. Archives of biochemistry and biophysics 330(1), 24-32,
Thienpont, B., Tingaud-Sequeira, A., Prats, E., Barata, C., Babin, P.J., Raldua, D., 2011. Zebrafish Eleutheroembryos Provide a Suitable Vertebrate Model for Screening Chemicals that Impair Thyroid Hormone Synthesis. Environmental Science & Technology 45, 7525-7532.
Tietge JE, Butterworth BC, Haselman JT, Holcombe GW, Hornung MW, Korte JJ, Kosian PA, Wolfe M, Degitz SJ. Early temporal effects of three thyroid hormone synthesis inhibitors in Xenopus laevis. Aquat Toxicol. 2010 98(1):44-50
van der Ven LTM, van den Brandhof EJ, Vos JH, Power DM, Wester PW. 2006. Effects of the antithyroid agent propylthiouracil in a partial life cycle assay with zebrafish. Environmental Science & Technology. 40(1):74-81.
Vickers AE, Heale J, Sinclair JR, Morris S, Rowe JM, Fisher RL. Thyroid organotypic rat and human cultures used to investigate drug effects on thyroid function, hormone synthesis and release pathways. Toxicol Appl Pharmacol. 2012 260(1):81-8.
Wabukebunoti MAN, Firling CE. 1983. The prehatching development of the thyroid-gland of the fathead minnow, pimephales-promelas (rafinesque). General and Comparative Endocrinology. 49(2):320-331.
Walter KM, Miller GW, Chen XP, Yaghoobi B, Puschner B, Lein PJ. 2019. Effects of thyroid hormone disruption on the ontogenetic expression of thyroid hormone signaling genes in developing zebrafish (danio rerio). General and Comparative Endocrinology. 272:20-32.
Walter, K.M., Miller, G.W., Chen, X.P., Yaghoobi, B., Puschner, B., Lein, P.J., 2019. Effects of thyroid hormone disruption on the ontogenetic expression of thyroid hormone signaling genes in developing zebrafish (Danio rerio). General and Comparative Endocrinology 272, 20-32.
Webster GM, Venners SA, Mattman A, Martin JW. 2014. Associations between perfluoroalkyl acids (pfass) and maternal thyroid hormones in early pregnancy: A population-based cohort study. Environmental Research. 133:338-347.
Relationship: 305: TH synthesis, Decreased leads to T4 in serum, Decreased
AOPs Referencing Relationship
Evidence Supporting Applicability of this Relationship
| Life Stage | Evidence |
|---|---|
| All life stages | High |
| Sex | Evidence |
|---|---|
| Male | High |
| Female | High |
Taxonomic: This KER is plausibly applicable across vertebrates. While a majority of the empirical evidence comes from work with laboratory rodents, there is a large amount of supporting data from humans (with anti-hyperthyroidism drugs including propylthiouracil and methimazole), some amphibian species (e.g., frog), fish species (e.g., zebrafish and fathead minnow), and some avian species (e.g, chicken). The following are samples from a large literature that supports this concept: Cooper et al. (1982; 1983); Hornung et al. (2010); Van Herck et al. (2013); Paul et al. (2013); Nelson et al. (2016); Alexander et al. (2017); Stinckens et al. (2020).
Life stage: Applicability to certain life stages may depend on the species and their dependence on maternally transferred thyroid hormones during the earliest phases of development. The earliest life stages of teleost fish rely on maternally transferred THs to regulate certain developmental processes until embryonic TH synthesis is active (Power et al., 2001). As a result, TPO inhibition is not expected to decrease TH synthesis during these earliest stages of development. In zebrafish, Opitz et al. (2011) showed the formation of a first thyroid follicle at 55 hours post fertilization (hpf), Chang et al. (2012) showed a first significant TH increase at 120 hpf and Walter et al. (2019) showed clear TH production already at 72 hpf but did not analyse time points between 24 and 72 hpf. In fathead minnows, a significant increase of whole body thyroid hormone levels was already observed between 1 and 2 dpf, which corresponds to the appearance of the thyroid anlage at 35 hpf prior to the first observation of thyroid follicles at 58 hpf (Wabuke-Bunoti and Firling, 1983). It is still uncertain when exactly embryonic TH synthesis is activated and how this determines sensitivity to TH disruptors.
Sex: The KE is plausibly applicable to both sexes. Thyroid hormones are essential in both sexes and the components of the HPT-axis are identical in both sexes. There can however be sex-dependent differences in the sensitivity to the disruption of thyroid hormone levels and the magnitude of the response. In humans, females appear more susceptible to hypothyroidism compared to males when exposed to certain halogenated chemicals (Hernandez‐Mariano et al., 2017; Webster et al., 2014). In adult zebrafish, Liu et al. (2019) showed sex-dependent changes in thyroid hormone levels and mRNA expression of regulatory genes including corticotropin releasing hormone (crh), thyroid stimulating hormone (tsh) and deiodinase 2 after exposure to organophosphate flame retardants. The underlying mechanism of any sex-related differences remains unclear.
Key Event Relationship Description
Thyroid hormones (THs), thyroxine (T4) and triiodothyronine (T3) are synthesized by NIS and TPO in the thyroid gland as iodinated thyroglobulin (Tg) and stored in the colloid of thyroid follicles across vertebrates. Secretion from the follicle into serum is a multi-step process. The first involves thyroid stimulating hormone (TSH) stimulation of the separation of the peptide linkage between Tg and TH. The next steps involve endocytosis of colloid, fusion of the endosome with the basolateral membrane of the thyrocyte, and finally release of TH into blood. More detailed descriptions of this process can be found in reviews by Braverman and Utiger (2012) and Zoeller et al. (2007).
Evidence Supporting this KER
The weight of evidence linking these two KEs of decreased TH synthesis and decreased T4 in serum is strong. It is commonly accepted dogma that decreased synthesis in the thyroid gland will result in decreased circulating TH (serum T4).
Biological PlausibilityThe biological relationship between two KEs in this KER is well understood and documented fact within the scientific community.
Empirical EvidenceIt is widely accepted that TPO inhibition leads to declines in serum T4 levels in adult mammals. This is due to the fact that the sole source for circulating T4 derives from hormone synthesis in the thyroid gland. Indeed, it has been known for decades that insufficient dietary iodine will lead to decreased serum TH concentrations due to inadequate synthesis. Strong qualitative and quantitative relationships exist between reduced TH synthesis and reduced serum T4 (Ekerot et al., 2013; Degon et al., 2008; Cooper et al., 1982; 1983; Leonard et al., 2016; Zoeller and Tan, 2007). There is more limited evidence supporting the relationship between decreased TH synthesis and lowered circulating hormone levels during development. Lu and Anderson (1994) followed the time course of TH synthesis, measured as thyroxine secretion rate, in non-treated pregnant rats and correlated it with serum T4 levels More recently, modeling of TH in the rat fetus demonstrates the quantitative relationship between TH synthesis and serum T4 concentrations (Hassan et al., 2017). Furthermore, a wide variety of drugs and chemicals that inhibit TPO are known to result in decreased release of TH from the thyroid gland, as well as decreased circulating TH concentrations. This is evidenced by a very large number of studies that employed a wide variety of techniques, including thyroid gland explant cultures, tracing organification of 131-I and in vivo treatment of a variety of animal species with known TPO inhibitors (King and May,1984; Atterwill et al., 1990; Brown et al., 1986; Brucker-Davis, 1998; Haselman et al., 2020; Hornung et al., 2010; Hurley et al., 1998; Kohrle, 2008; Tietge et al., 2010).
Additionally, evidence is available from studies investigating responses to TPO inhibitors in fish. For example, Stinckens et al. (2020) showed reduced whole body T4 concentrations in zebrafish larvae exposed to 50 or 100 mg/L methimazole, a potent TPO inhibitor, from immediately after fertilization until 21 or 32 days of age. Exposure to 37 or 111 mg/L propylthiouracil also reduced T4 levels after exposure up to 14, 21 and 32 days in the same study. Walter et al. (2019) showed that propylthiouracil had no effect on T4 levels in 24h old zebrafish, but decreased T4 levels of 72h old zebrafish. This difference is probably due to the onset of embryonic TH production between the age of 24 and 72 hours (Opitz et al., 2011). Stinckens et al. (2016) showed that exposure to 2-mercaptobenzothiazole (MBT), an environmentally relevant TPO inhibitor, decreased whole body T4 levels in continuously exposed 5 and 32 day old zebrafish larvae. Several other studies have also shown that chemically induced Inhibition of TPO results in reduced TH synthesis in zebrafish (Van der Ven et al., 2006; Raldua and Babin, 2009; Liu et al., 2011; Thienpont et al., 2011; Rehberger et al., 2018). A high concentration of MBT also decreased whole body T4 levels in 6 day old fathead minnows, but recovery was observed at the age of 21 days although the fish were kept in the exposure medium (Nelson et al., 2016). Crane et al. (2006) showed decreased T4 levels in 28 day old fathead minnows continuously exposed to 32 or 100 µg/L methimazole.
Temporal Evidence: In mammals, the temporal nature of this KER is applicable to all life stages, including development (Seed et al., 2005). There are currently no studies that measured both TPO synthesis and TH production during development. However, the impact of decreased TH synthesis on serum hormones is similar across all ages in mammals. Good evidence for the temporal relationship comes from thyroid system modeling of the impacts of iodine deficiency and NIS inhibition (e.g., Degon et al., 2008; Fisher et al., 2013). In addition, recovery experiments have demonstrated that serum thyroid hormones recovered in athyroid mice following grafting of in-vitro derived follicles (Antonica et al., 2012). In Xenopus, it has been shown that depression of TH synthesis in the thyroid gland precedes depression of circulating TH within 7 days of exposure during pro-metamorphosis (Haselman et al., 2020).
In oviparous fish such as zebrafish and fathead minnow, the nature of this KER depends on the life stage since the earliest stages of embryonic development rely on maternal thyroid hormones transferred to the eggs. Embryonic thyroid hormone synthesis is activated later during embryo-larval development. (See Domain of applicability)
Dose-response Evidence: Dose-response data is lacking from studies that include concurrent measures of both TH synthesis and serum TH concentrations. However, data is available demonstrating correlations between thyroidal TH and serum TH concentrations during gestation and lactation during development (Gilbert et al., 2013). This data was used to develop a rat quantitative biologically-based dose-response model for iodine deficiency (Fisher et al., 2013). In Xenopus, dose-responses were demonstrated in both thyroidal T4 and circulating T4 following exposure to three TPO inhibitors (Haselman et al., 2020).
Uncertainties and InconsistenciesThere are no inconsistencies in this KER, but there are some uncertainties. The first uncertainty stems from the paucity of data for quantitative modeling of the relationship between the degree of synthesis decrease and resulting changes in circulating T4 concentrations. In addition, most of the data supporting this KER comes from inhibition of TPO, and there are a number of other processes (e.g., endocytosis, lysosomal fusion, basolateral fusion and release) that are not as well studied.
For example, Kim et al. (2015) investigated the adverse effects of Triphenyl phosphate (TPP), a substance that disrupts the thyroid system.
Therefore, Rat pituitary (GH3) and thyroid follicular cell lines (FRTL-5) were studied. In the GH3 cells, TPP led to an upregulation of the expression of important thyroid genes (tshˇ, tr˛ and trˇ) while T3, a positive control, downregulated the expression of these genes. In FRTL-5 cells, the expression of nis and tpo genes was significantly upregulated,
suggesting that TPP stimulates thyroid hormone synthesis in the thyroid gland.
In zebrafish larvae at the age of 7 days post-fertilisation (dpf), TPP exposure resulted in a significant increase in T3 and T4 concentrations and the expression of genes involved in thyroid hormone synthesis. Exposure to TPP also significantly regulated the expression of genes involved in the metabolism (dio1), transport (ttr) and excretion
(ugt1ab) of thyroid hormones. The down-regulation of the crh and tshˇ genes in the zebrafish larvae
suggests the activation of a central regulatory feedback mechanism that is triggered by the increased T3 levels in
vivo. Taken together, these observations indicate that TPP increases thyroid hormone concentrations
in early life stages of zebrafish by disrupting central regulatory and hormone synthesis pathways.
Quantitative Understanding of the Linkage
In rats, Hassan et al. (2020) demonstrated in vitro:ex vivo correlations of TPO inhibition using PTU and MMI and constructed a quantitative model relating level of TPO inhibition with changes in circulating T4 levels. They determined that 30% inhibition of TPO was sufficient to decrease circulating T4 levels by 20%.
In Xenopus, Haselman et al. (2020) collected temporal and dose-response data for both thyroidal and circulating T4 which showed strong qualitative concordance of the response-response relationship. A quantitative relationship exists therein, but is yet to be demonstrated mathematically in this species.
Response-response relationshipFisher et al. (2013) published a quantitative biologically-based dose-response model for iodine deficiency in the rat. This model provides quantitative relationships for thyroidal T4 synthesis (iodine organification) and predictions of serum T4 concentrations in developing rats. There are other computational models that include thyroid hormone synthesis. Ekerot et al. (2012) modeled TPO, T3, T4 and TSH in dogs and humans based on exposure to myeloperoxidase inhibitors that also inhibit TPO. This model was recently adapted for rat (Leonard et al., 2016) and Hassan et al (2017) have extended it to include the pregnant rat dam in response to TPO inhibition induced by PTU. While the original model predicted serum TH and TSH levels as a function of oral dose, it was not used to explicitly predict the relationship between serum hormones and TPO inhibition, or thyroidal hormone synthesis. Leonard et al. (2016) recently incorporated TPO inhibition into the model. Degon et al (2008) developed a human thyroid model that includes TPO, but does not make quantitative prediction of organification changes due to inhibition of the TPO enzyme. Further empirical support for the response-response relationship has been demonstrated in the amphibian model, Xenopus laevis, exposed to TPO inhibitors during pro-metamorphosis (Haselman et al., 2020) wherein temporal profiles were measured for both thyroidal and circulating T4.
Time-scaleGiven that the thyroid gland contains follicular lumen space filled with stored thyroglobulin/T4, complete inhibition of thyroid hormone synthesis at a given point in time will not result in an instantaneous decrease in circulating T4. The system will be capable of maintaining sufficient circulating T4 levels until the gland stores are depleted. The time it takes to deplete stored hormone will greatly depend on species, developmental status and numerous other factors.
In Xenopus, Haselman et al. (2020) demonstrated an approximately 5 day difference between a significant decrease in thyroidal T4 preceding a significant decrease in circulating T4 while exposed to a potent TPO inhibitor (MMI) continuously during pro-metamorphosis.
Known modulating factorsDuring Xenopus metamorphosis, circulating T4 steadily increases to peak levels at metamorphic climax. Therefore, during Xenopus metamorphosis, this KER is operable at an increased rate as compared to a system that is maintaining steady circulating T4 levels through homeostatic control. In this case, developmental status is a modulating factor for the rates and trajectories of these KEs.
Known Feedforward/Feedback loops influencing this KERThis KER is entirely influenced by the feedback loop between circulating T4 originating from the thyroid gland and circulating TSH originating from the pituitary. Intermediate biochemical processes exist within the hypothalamus to affirm feedback and coordinately release TSH from the pituitary. However, quantitative representations of these feedback processes are limited to models discussed previously.
In Xenopus, circulating levels of T4 increase through pro-metamorphosis indicating a "release" of feedback to allow circulating levels of T4 to increase and drive metamorphic changes (Sternberg et al., 2011). This provides evidence that homeostatic control of feedback can be developmentally dependent, and likely species dependent.
References
Alexander EK, Pearce EN, Brent GA, Brown RS, Chen H, Dosiou C, Grobman WA, Laurberg P, Lazarus JH, Mandel SJ, Peeters RP, Sullivan S. 2017 Guidelines of the American Thyroid Association for the Diagnosis and Management of Thyroid Disease During Pregnancy and the Postpartum. Thyroid. 2017 Mar;27(3):315-389.
Antonica F, Kasprzyk DF, Opitz R, Iacovino M, Liao XH, Dumitrescu AM, Refetoff S, Peremans K, Manto M, Kyba M, Costagliola S. Generation of functional thyroid from embryonic stem cells. Nature. 2012 491(7422):66-71.
Atterwill CK, Fowler KF. A comparison of cultured rat FRTL-5 and porcine thyroid cells for predicting the thyroid toxicity of xenobiotics. Toxicol In Vitro. 1990. 4(4-5):369-74.
Braverman, L.E. and Utiger, R.D. (2012). Werner and Ingbar's The Thyroid: A Fundamental and Clinical Text (10 ed.). Philadelphia, PA: Lippincott Williams & Wilkins. pp. 775-786. ISBN 978-1451120639.
Brown CG, Fowler KL, Nicholls PJ, Atterwill C. Assessment of thyrotoxicity using in vitro cell culture systems. Food Chem Toxicol. 1986 24(6-7):557-62.
Brucker-Davis F. Effects of environmental synthetic chemicals on thyroid function. Thyroid. 1998 8(9):827-56.
Chang J, Wang M, Gui W, Zhao Y, Yu L, Zhu G. 2012. Changes in thyroid hormone levels during zebrafish development. Zoological Science. 29(3):181-184.
Cooper DS, Kieffer JD, Halpern R, Saxe V, Mover H, Maloof F, Ridgway EC (1983) Propylthiouracil (PTU) pharmacology in the rat. II. Effects of PTU on thyroid function. Endocrinology 113:921-928.
Cooper DS, Saxe VC, Meskell M, Maloof F, Ridgway EC.Acute effects of propylthiouracil (PTU) on thyroidal iodide organification and peripheral iodothyronine deiodination: correlation with serum PTU levels measured by radioimmunoassay. J Clin Endocrinol Metab. 1982 54(1):101-7.
Crane HM, Pickford DB, Hutchinson TH, Brown JA. 2006. The effects of methimazole on development of the fathead minnow, pimephales promelas, from embryo to adult. Toxicological Sciences. 93(2):278-285.
Degon, M., Chipkin, S.R., Hollot, C.V., Zoeller, R.T., and Chait, Y. (2008). A computational model of the human thyroid. Mathematical Biosciences 212, 22–53
Ekerot P, Ferguson D, Glämsta EL, Nilsson LB, Andersson H, Rosqvist S, Visser SA. Systems pharmacology modeling of drug-induced modulation of thyroid hormones in dogs and translation to human. Pharm Res. 2013 30(6):1513-24.
Fisher JW, Li S, Crofton K, Zoeller RT, McLanahan ED, Lumen A, Gilbert ME. Evaluation of iodide deficiency in the lactating rat and pup using a biologically based dose-response model. Toxicol Sci. 2013 132(1):75-86.
Gilbert ME, Hedge JM, Valentín-Blasini L, Blount BC, Kannan K, Tietge J, Zoeller RT, Crofton KM, Jarrett JM, Fisher JW. An animal model of marginal iodine deficiency during development: the thyroid axis and neurodevelopmental outcome. Toxicol Sci. 2013 132(1):177-95.
Haselman, J.T., Olker, J.H., Kosian, P.A., Korte, J.J., Swintek, J.A., Denny, J.S., Nichols, J.W., Tietge, J.E., Hornung, M.W. and Degitz, S.J., 2020. Targeted pathway-based in vivo testing using thyroperoxidase inhibition to evaluate plasma thyroxine as a surrogate metric of metamorphic success in model amphibian Xenopus laevis. Toxicological Sciences, 175(2), pp.236-250.
Hassan, I, El-Masri, H., Kosian, PA, Ford, J, Degitz, SJ and Gilbert, ME. Quantitative Adverse Outcome Pathway for Neurodevelopmental Effects of Thyroid Peroxidase-Induced Thyroid Hormone Synthesis Inhibition. Toxicol Sci. 2017 Nov 1;160(1):57-73
Hassan, I., El-Masri, H., Ford, J., Brennan, A., Handa, S., Paul Friedman, K. and Gilbert, M.E., 2020. Extrapolating in vitro screening assay data for thyroperoxidase inhibition to predict serum thyroid hormones in the rat. Toxicological Sciences, 173(2), pp.280-292.
Hernandez-Mariano JA, Torres-Sanchez L, Bassol-Mayagoitia S, Escamilla-Nunez M, Cebrian ME, Villeda-Gutierrez EA, Lopez-Rodriguez G, Felix-Arellano EE, Blanco-Munoz J. 2017. Effect of exposure to p,p '-dde during the first half of pregnancy in the maternal thyroid profile of female residents in a mexican floriculture area. Environmental Research. 156:597-604.
Hornung MW, Degitz SJ, Korte LM, Olson JM, Kosian PA, Linnum AL, Tietge JE. Inhibition of thyroid hormone release from cultured amphibian thyroid glands by methimazole, 6-propylthiouracil, and perchlorate. Toxicol Sci. 2010 118(1):42-51.
Hurley PM. Mode of carcinogenic action of pesticides inducing thyroid follicular cell tumors in rodents. Environ Health Perspect. 1998 106(8):437-45.
Kim, S., Jung, J., Lee, I., Jung, D., Youn, H., & Choi, K. (2015). Thyroid disruption by triphenyl phosphate, an organophosphate flame retardant, in zebrafish (Danio rerio) embryos/larvae, and in GH3 and FRTL-5 cell lines. Aquatic Toxicology, 160, 188–196. https://doi.org/10.1016/j.aquatox.2015.01.016
King DB, May JD. Thyroidal influence on body growth. J Exp Zool. 1984 Dec;232(3):453-60.
Köhrle J. Environment and endocrinology: the case of thyroidology. Ann Endocrinol (Paris). 2008 69(2):116-22.
Leonard JA, Tan YM, Gilbert M, Isaacs K, El-Masri H. Estimating margin of exposure to thyroid peroxidase inhibitors using high-throughput in vitro data, high-throughput exposure modeling, and physiologically based pharmacokinetic/pharmacodynamic modeling. Toxicol Sci. 2016 151(1):57-70.
Liu CS, Zhang XW, Deng J, Hecker M, Al-Khedhairy A, Giesy JP, Zhou BS. 2011. Effects of prochloraz or propylthiouracil on the cross-talk between the hpg, hpa, and hpt axes in zebrafish. Environmental Science & Technology. 45(2):769-775.
Liu XS, Cai Y, Wang Y, Xu SH, Ji K, Choi K. 2019. Effects of tris(1,3-dichloro-2-propyl) phosphate (tdcpp) and triphenyl phosphate (tpp) on sex-dependent alterations of thyroid hormones in adult zebrafish. Ecotoxicology and Environmental Safety. 170:25-32.
Lu, M-H, and Anderson, RR. Thyroxine secretion rats during pregnancy in the rat. Endo Res. 1994. 20(4):343-364.
Nelson K, Schroeder A, Ankley G, Blackwell B, Blanksma C, Degitz S, Flynn K, Jensen K, Johnson R, Kahl M et al. 2016. Impaired anterior swim bladder inflation following exposure to the thyroid peroxidase inhibitor 2-mercaptobenzothiazole part i: Fathead minnow. Aquatic Toxicology. 173:192-203.
Opitz R, Maquet E, Zoenen M, Dadhich R, Costagliola S. 2011. Tsh receptor function is required for normal thyroid differentiation in zebrafish. Molecular Endocrinology. 25(9):1579-1599.
Paul KB, Hedge JM, Macherla C, Filer DL, Burgess E, Simmons SO, Crofton KM, Hornung MW. Cross-species analysis of thyroperoxidase inhibition by xenobiotics demonstrates conservation of response between pig and rat. Toxicology. 2013. 312:97-107.
Power DM, Llewellyn L, Faustino M, Nowell MA, Bjornsson BT, Einarsdottir IE, Canario AV, Sweeney GE. 2001. Thyroid hormones in growth and development of fish. Comp Biochem Physiol C Toxicol Pharmacol. 130(4):447-459.
Raldua D, Babin PJ. 2009. Simple, rapid zebrafish larva bioassay for assessing the potential of chemical pollutants and drugs to disrupt thyroid gland function. Environmental Science & Technology. 43(17):6844-6850.
Rehberger K, Baumann L, Hecker M, Braunbeck T. 2018. Intrafollicular thyroid hormone staining in whole-mount zebrafish (danio rerio) embryos for the detection of thyroid hormone synthesis disruption. Fish Physiology and Biochemistry. 44(3):997-1010.
Sternberg, R.M., Thoemke, K.R., Korte, J.J., Moen, S.M., Olson, J.M., Korte, L., Tietge, J.E. and Degitz Jr, S.J., 2011. Control of pituitary thyroid-stimulating hormone synthesis and secretion by thyroid hormones during Xenopus metamorphosis. General and comparative endocrinology, 173(3), pp.428-437.
Stinckens E, Vergauwen L, Blackwell BR, Anldey GT, Villeneuve DL, Knapen D. 2020. Effect of thyroperoxidase and deiodinase inhibition on anterior swim bladder inflation in the zebrafish. Environmental Science & Technology. 54(10):6213-6223.
Stinckens E, Vergauwen L, Schroeder A, Maho W, Blackwell B, Witters H, Blust R, Ankley G, Covaci A, Villeneuve D et al. 2016. Impaired anterior swim bladder inflation following exposure to the thyroid peroxidase inhibitor 2-mercaptobenzothiazole part ii: Zebrafish. Aquatic Toxicology. 173:204-217.
Thienpont B, Tingaud-Sequeira A, Prats E, Barata C, Babin PJ, Raldua D. 2011. Zebrafish eleutheroembryos provide a suitable vertebrate model for screening chemicals that impair thyroid hormone synthesis. Environmental Science & Technology. 45(17):7525-7532.
Tietge, J.E., Butterworth, B.C., Haselman, J.T., Holcombe, G.W., Hornung, M.W., Korte, J.J., Kosian, P.A., Wolfe, M. and Degitz, S.J., 2010. Early temporal effects of three thyroid hormone synthesis inhibitors in Xenopus laevis. Aquatic Toxicology, 98(1), pp.44-50.
van der Ven LTM, van den Brandhof EJ, Vos JH, Power DM, Wester PW. 2006. Effects of the antithyroid agent propylthiouracil in a partial life cycle assay with zebrafish. Environmental Science & Technology. 40(1):74-81.
Van Herck SL, Geysens S, Delbaere J, Darras VM. Regulators of thyroid hormone availability and action in embryonic chicken brain development. Gen Comp Endocrinol. 2013. 190:96-104.
Wabukebunoti MAN, Firling CE. 1983. The prehatching development of the thyroid-gland of the fathead minnow, pimephales-promelas (rafinesque). General and Comparative Endocrinology. 49(2):320-331.
Walter KM, Miller GW, Chen XP, Yaghoobi B, Puschner B, Lein PJ. 2019. Effects of thyroid hormone disruption on the ontogenetic expression of thyroid hormone signaling genes in developing zebrafish (danio rerio). General and Comparative Endocrinology. 272:20-32.
Webster GM, Venners SA, Mattman A, Martin JW. 2014. Associations between perfluoroalkyl acids (pfass) and maternal thyroid hormones in early pregnancy: A population-based cohort study. Environmental Research. 133:338-347.
Zoeller, R. T., Tan, S. W., and Tyl, R. W. (2007). General background on the hypothalamic-pituitary-thyroid (HPT) axis. Critical reviews in toxicology 37(1-2), 11-53.
Relationship: 2038: T4 in serum, Decreased leads to Decreased, Triiodothyronine (T3) in serum
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Thyroperoxidase inhibition leading to increased mortality via reduced anterior swim bladder inflation | adjacent | Moderate | Moderate |
| Thyroperoxidase inhibition leading to altered visual function via altered retinal layer structure | adjacent | Moderate | Moderate |
| Thyroperoxidase inhibition leading to altered visual function via decreased eye size | adjacent | ||
| Thyroperoxidase inhibition leading to altered visual function via altered photoreceptor patterning | adjacent |
Evidence Supporting Applicability of this Relationship
| Life Stage | Evidence |
|---|---|
| Juvenile | Moderate |
| Larvae | Moderate |
| Sex | Evidence |
|---|---|
| Unspecific | Moderate |
Taxonomic: Thyroid follicles mainly produce T4 and to a lesser extent T3 across vertebrates. When serum T4 levels are decreased, less T4 is available for conversion to the more biologically active T3. This key event relationship is not always evident. This could be due to feedback/compensatory mechanisms that in some cases seem to be able to maintain T3 levels even though T4 levels are reduced, for example through increased conversion of T4 to T3 by deiodinases. These feedback mechanisms can also differ across species. Therefore, although this KER is plausibly applicable across vertebrates, variation can be expected. In zebrafish and fathead minnow, several studies reportedThe evidence for a relationship between circulating T4 and T3 levels (Nelson et al., 2016; Stinckens et al., 2020, Wang et al., 2020).
Life stage: This key event relationship is applicable to late larvae and juveniles rather than to embryos, because of the presence of maternal TH in embryos.
Uncertainties during embryonic lifestage:
- A decrease in T4 was observed in fathead minnows exposed to 1 mg/L 2-mercaptobenzothiazole (MBT), a thyroperoxidase inhibitor, through 6 dpf (Nelson et al., 2016). In contrast, there was no observed effect on T3 in fathead minnows exposed to MBT through 6 dpf. Comparably, zebrafish exposed to 0.4 or 0.7 mg/L MBT through 120 hpf showed decreased T4 but not T3 (Stinckens et al., 2016). During this early larval life stage, T3 may have been derived from maternal T4. In addition, it could be produced from further depletion of any T4 still produced by the thyroid gland (as thyroperoxidase may not have been fully inhibited at the tested exposure concentrations).
- Since exposure to PFAS did result in decreased whole-body T4 and T3 in 5 day old zebrafish, the life-stage specificity possibly depends on the mechanism that lies at the basis of the TH changes (Wang et al., 2020). The exact mechanisms by which PFAS disrupt the thyroid hormone system remain uncertain. Compounds that directly reduce T3 levels (e.g., deiodinase inhibitors) in addition to reducing T4 levels via another mechanism can be expected to result in decreased T4 and T3 levels.
Sex: The KE is plausibly applicable to both sexes. Thyroid hormones are essential in both sexes and the components of the HPT-axis are identical in both sexes. There can however be sex-dependent differences in the sensitivity to the disruption of thyroid hormone levels and the magnitude of the response. In humans, females appear more susceptible to hypothyroidism compared to males when exposed to certain halogenated chemicals (Hernandez‐Mariano et al., 2017; Webster et al., 2014). In adult zebrafish, Liu et al. (2019) showed sex-dependent changes in thyroid hormone levels and mRNA expression of regulatory genes including corticotropin releasing hormone (crh), thyroid stimulating hormone (tsh) and deiodinase 2 after exposure to organophosphate flame retardants. The underlying mechanism of any sex-related differences remains unclear.
Key Event Relationship Description
When serum thyroxine (T4) levels are decreased, less T4 is available for conversion to the more biologically active triiodothyronine (T3). While some thyroid hormone (TH) disrupting mechanisms can immediately affect T3 levels, including deiodinase inhibition, other mechanisms reduce T4 levels, for example through inhibition of TH synthesis, leading to decreased T3 levels.
Since in fish early life stages TH are typically measured on a whole body level, it is currently uncertain whether TH levels changes occur at the serum and/or tissue level. Pending more dedicated studies, whole body TH levels are considered a proxy for serum TH levels.
This key event relationship is not always evident. This could be due to feedback/compensatory mechanisms that in some cases seem to be able to maintain T3 levels even though T4 levels are reduced, for example through increased conversion of T4 to T3 by deiodinases.
Evidence Supporting this KER
Biological PlausibilityWhen serum thyroxine (T4) levels are decreased, less T4 is available for conversion to the more biologically active triiodothyronine (T3). It is plausible to assume that while some thyroid hormone (TH) disrupting mechanisms can immediately affect T3 levels, including deiodinase inhibition, other mechanisms reduce T4 levels, for example through inhibition of TH synthesis, leading to decreased T3 levels.
Empirical Evidence- A decrease in whole-body T4 and T3 was observed in zebrafish exposed to methimazole from fertilization until the age of 21 and 32 days and to propylthiouracil until the age of 14, 21 and 32 days (Stinckens et al., 2020). Additionally, a strong correlation was observed between T4 and T3 levels. Both compounds are thyroperoxidase inhibitors expected to inhibit thyroid hormone synthesis.
- A dose-dependent decrease in whole-body T4 and T3 was observed in zebrafish exposed to perfluorooctanoic acid and perfluoropolyether carboxylic acids from fertilization until the age of 5 days (Wang et al., 2020). The exact mechanisms by which PFAS disrupt the thyroid hormone system remain uncertain.
- While T4 measurements could not be acquired in fathead minnows exposed to 1 mg/L 2-mercaptobenzothiazole, a thyroperoxidase inhibitor, for 14 days, a significant decrease in T3 was observed (Nelson et al., 2016). The decreased T3 levels were likely the result of reduced T4 synthesis.
- Since in fish early life stages THs are typically measured on a whole body level, it is currently uncertain whether TH level changes occur at the serum and/or tissue level. Pending more dedicated studies, whole body TH levels are considered a proxy for serum TH levels.
- This key event relationship is not always evident. This could be due to feedback/compensatory mechanisms that in some cases seem to be able to maintain T3 levels even though T4 levels are reduced, for example through increased conversion of T4 to T3 by deiodinases. Examples of studies showing reduced T4 levels in the absence of reduced T3 levels:
- Zebrafish exposed to 0.35 mg/L 2-mercaptobenzothiazole, a thyroperoxidase inhibitor, through 32 dpf showed decreased whole-body T4, but T3 levels showed particularly large variation and overall were not significantly decreased (Stinckens et al., 2016).
- Although T4 content of 28 dpf larval fathead minnows exposed to 32 or 100 µg/l methimazole, a thyroperoxidase inhibitor, was reduced, these fish showed no change in whole body T3 content (Crane et al., 2006). Significantly higher T3/T4 ratios in fish held in 100 µg/l methimazole suggest an increased conversion of T4 to T3 or reduced degradation and conjugation during continued exposure to methimazole
Quantitative Understanding of the Linkage
Stinckens et al. (2020, supplementary information) showed a significant linear relationship between whole body T3 and T4 concentrations at 21 and 32 days post fertilization after continuous exposure of zebrafish to methimazole and propylthiouracil, two inhibitors of TH synthesis.
Known Feedforward/Feedback loops influencing this KERThis key event relationship is not always evident. This could be due to feedback/compensatory mechanisms that in some cases seem to be able to maintain T3 levels even though T4 levels are reduced, for example through increased conversion of T4 to T3 by deiodinases. Examples of studies showing reduced T4 levels in the absence of reduced T3 levels:
- Zebrafish exposed to 0.35 mg/L 2-mercaptobenzothiazole, a thyroperoxidase inhibitor, through 32 dpf showed decreased whole-body T4, but T3 levels showed particularly large variation and overall were not significantly decreased (Stinckens et al., 2016).
- Although T4 content of 28 dpf larval fathead minnows exposed to 32 or 100 µg/l methimazole, a thyroperoxidase inhibitor, was reduced, these fish showed no change in whole body T3 content (Crane et al., 2006). Significantly higher T3/T4 ratios in fish held in 100 µg/l methimazole suggest an increased conversion of T4 to T3 or reduced degradation and conjugation during continued exposure to methimazole
This relationship depends on the MIE that is causing the decrease in T3. For example, deiodinase inhibition results in reduced activation of T4 to T3 and thus in reduced T3 levels; increased T4 levels have been observed, probably as a compensatory mechanism in response to the lower T3 levels. For example, Cavallin et al. (2017) exposed fathead minnows to iopanoic acid, a deiodinase inhibitor, and observed T4 increases together with T3 decreases.
References
Cavallin, J.E., Ankley, G.T., Blackwell, B.R., Blanksma, C.A., Fay, K.A., Jensen, K.M., Kahl, M.D., Knapen, D., Kosian, P.A., Poole, S.T., Randolph, E.C., Schroeder, A.L., Vergauwen, L., Villeneuve, D.L., 2017. Impaired swim bladder inflation in early life stage fathead minnows exposed to a deiodinase inhibitor, iopanoic acid. Environmental Toxicology and Chemistry 36, 2942-2952.
Crane, H.M., Pickford, D.B., Hutchinson, T.H., Brown, J.A., 2006. The effects of methimazole on development of the fathead minnow, pimephales promelas, from embryo to adult. Toxicological Sciences 93, 278-285.
Hernandez-Mariano JA, Torres-Sanchez L, Bassol-Mayagoitia S, Escamilla-Nunez M, Cebrian ME, Villeda-Gutierrez EA, Lopez-Rodriguez G, Felix-Arellano EE, Blanco-Munoz J. 2017. Effect of exposure to p,p '-dde during the first half of pregnancy in the maternal thyroid profile of female residents in a mexican floriculture area. Environmental Research. 156:597-604.
Liu XS, Cai Y, Wang Y, Xu SH, Ji K, Choi K. 2019. Effects of tris(1,3-dichloro-2-propyl) phosphate (tdcpp) and triphenyl phosphate (tpp) on sex-dependent alterations of thyroid hormones in adult zebrafish. Ecotoxicology and Environmental Safety. 170:25-32.
Nelson, K., Schroeder, A., Ankley, G., Blackwell, B., Blanksma, C., Degitz, S., Flynn, K., Jensen, K., Johnson, R., Kahl, M., Knapen, D., Kosian, P., Milsk, R., Randolph, E., Saari, T., Stinckens, E., Vergauwen, L., Villeneuve, D., 2016. Impaired anterior swim bladder inflation following exposure to the thyroid peroxidase inhibitor 2-mercaptobenzothiazole part I: Fathead minnow. Aquatic Toxicology 173, 192-203.
Stinckens, E., Vergauwen, L., Blackwell, B.R., Anldey, G.T., Villeneuve, D.L., Knapen, D., 2020. Effect of Thyroperoxidase and Deiodinase Inhibition on Anterior Swim Bladder Inflation in the Zebrafish. Environmental Science & Technology 54, 6213-6223.
Stinckens, E., Vergauwen, L., Schroeder, A., Maho, W., Blackwell, B., Witters, H., Blust, R., Ankley, G., Covaci, A., Villeneuve, D., Knapen, D., 2016. Impaired anterior swim bladder inflation following exposure to the thyroid peroxidase inhibitor 2-mercaptobenzothiazole part II: Zebrafish. Aquatic Toxicology 173, 204-217.
Wang, J.X., Shi, G.H., Yao, J.Z., Sheng, N., Cui, R.N., Su, Z.B., Guo, Y., Dai, J.Y., 2020. Perfluoropolyether carboxylic acids (novel alternatives to PFOA) impair zebrafish posterior swim bladder development via thyroid hormone disruption. Environment International 134.
Webster GM, Venners SA, Mattman A, Martin JW. 2014. Associations between perfluoroalkyl acids (pfass) and maternal thyroid hormones in early pregnancy: A population-based cohort study. Environmental Research. 133:338-347.
Relationship: 2373: Decreased, Triiodothyronine (T3) in serum leads to Altered, retinal layer structure
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Thyroperoxidase inhibition leading to altered visual function via altered retinal layer structure | adjacent |
Evidence Supporting Applicability of this Relationship
| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| zebrafish | Danio rerio | NCBI |
| Life Stage | Evidence |
|---|---|
| Embryo | Moderate |
| Sex | Evidence |
|---|---|
| Unspecific |
Life-stage applicability: Most studies on TH-regulated retinal structure are performed during vertebrate development. There is evidence of the impact of reduced T3 (caused by inhibition of thyroperoxidase) on retinal layer structure at 48, 66, 72, 96 and 120 hpf during zebrafish embryo-eleutheroembryo development (Baumann and others 2016; Komoike and others 2013; Reider and Connaughton 2014).
Taxonomic applicability: The visual system of the zebrafish follows the typical organisation of vertebrates and is often used as a model to study human eye diseases. Although there are some differences in eye structure between zebrafish and humans, it is plausible to assume that TH levels are important for healthy eye development across all vertebrates.
Sex applicability: Zebrafish are undifferentiated gonochorists since both sexes initially develop an immature ovary (Maack and Segner, 2003). Immature ovary development progresses until approximately the onset of the third week. Later, in female fish immature ovaries continue to develop further, while male fish undergo transformation of ovaries into testes. Final transformation into testes varies among male individuals, however finishes usually around 6 weeks post fertilization. Effects on retinal layers resulting from TH level changes during early development are therefore expected to be independent of sex.
Key Event Relationship Description
Although the exact mechanisms need further investigation, studies show that thyroid hormones (THs) are required for healthy eye development in vertebrates (Wester et al. 1990, Suliman & Novales Flamarique 2013, Deveau et al., 2019) and it has been described that retinal development, photoreceptor differentiation and colour vision are directly regulated by THs. Not only in zebrafish (Bertrand et al. 2007), but also in mice (Ng et al. 2001) and chickens (Trimarchi et al. 2008), THs are directly linked to the transcription of essential visual opsins and the differentiation of retinal cells, as well as the overall structure of the retina, which is essential for proper functioning. Therefore, decreased triidothyronine (T3) levels in serum during eye development are likely to lead to structural and morphological alterations of the eyes.
Evidence Supporting this KER
Biological PlausibilityTHs, TH receptors, and deiodinase (DIO) enzymes are important for eye and retinal development in vertebrates. Dio enzymes activate and inactivate THs, consequently playing a central role in regulating TH levels in target tissues. In zebrafish, TH receptors and dio enzymes have been localized in the retina from 24 hpf onwards, probably regulating the differentiation of retinal structures and photoreceptors. It is known from amphibians that when TH levels start to rise at the beginning of metamorphosis, the morphology of the eyes starts changing. In chicken, the developing eye shows a dynamic expression pattern of Deiodinase 2 (DIO2) and Deiodinase 3 (DIO3), probably regulating photoreceptor differentiation and cornea development (reviewed by Darras 2015).
There is ample evidence across vertebrates that THs have an influence on development of the retinal layer structure:
-
Baumann (2016) described alterations in eye structure, size and pigmentation in PTU- (6-n-propylthouracil is a classic positive control for inhibition of TPO, reduced TH levels) and TBBPA- (TR activation) treated zebrafish embryos.
-
Komoike et al. (2013) exposed zebrafish embryos to 10 mM methimazole (a TPO inhibitor that is expected to result in lower TH levels) and observed moderately disrupted retinal structure with apoptosis of renital cells already at 48 hpf and more severely disrupted retinal structure at 72 hpf.
-
Besson et al. (2020) used pharmacological treatments (T3 + iopanoic acid (IOP), NH3, Chlorpyrifos (CPF)) to highlight the role that thyroid hormones (TH) play in sensory development in metamorphosing convict surgeonfish. They found out that co-exposure to +1.5 °C and CPF5 significantly decreased T4 and T3 levels (43% and 26%, respectively) and N3-treated fish experienced repressed sensory development, at both day 2 and day 5 for the retina. This study highlights that short-term exposure to acute anthropogenic stressors has adverse consequences for the development, behavior, and survival of metamorphosing fish by affecting their TH endocrine function.
-
Bhumika et al. (2014) found that lowering T3 signaling through exposure to different chemicals accelerates optic tectum reinnervation following optiv nerve crush in zebrafish and that this is accompanied by a more rapid resolution of the inflammatory response.
-
Using genetic experiments Duval and Allison (2018) showed that disruption of thyroid receptors as well as disruption of cone development factors such as gdf6a and tbx2b lead to altered photoreceptor structure.
-
Reider and Connaughton (2014) exposed zebrafish embryos to methimazole (MMI), a TPO inhibitor that is expected to result in lower TH levels, until 66, 70 or 72 hpf and analysed the retina at 72 hpf. They observed a reduced ganglion cell layer (GCL) diameter in embryos exposed until 66 hpf, but not in embryos exposed until 70 or 72 hpf. The thickness of the GCL was decreased in larvae exposed to MMI until 66 hpf compared to controls. An increase in GCL thickness was observed at 70 hpf, followed by a decrease in thickness at 72 hpf (p < 0.05). IPL thickness measurements also showed the smallest value in retinas exposed to MMI until 66 hpf.
-
Gamborino (2000) analysed eye development in a rat model of congenital-neonatal hypothyroidism (HG), induced by combined chemical-surgical thyroidectomy. Histopathological analyses of the eyes of TH-deficient animals revealed decrease in photoreceptor and ganglion cell layer thickness, a delay in photoreceptor outer segment morphogenesis and significantly lower values for ganglion cell nuclear volumes and nuclear pore density.
-
Gan et al. (2010) showed that thyroid hormones accelerate opsin expression in differentiating cones and induce the opsin
switch in differentiated single cones. The results show, that Propylthiouracil, a pharmacological inhibitor of thyroid hormone synthesis, repressed the opsin switch. Thyroid hormone is required for opsin switching in the retina of salmonid fishes. -
Houbrechts (2016) performed deiodinase knockdown in zebrafish embryos and observed reduced eye size, disturbed retinal lamination and strong reduction in rods and all four cone types. Defects were more prominent and persistent in D3-deficient fish, probably due to increased T3 signalling.
-
Wang et al. (2017) performed rescue experiments with the thyroid disruptors paclobutrazol (PBZ) and retinoic acid (RA). They showed that PBZ dramatically reduces photoreceptor proliferation. Co-treatment of PBZ with RA, or post-treatment of PBZ-treated embryos with RA, partially rescues photoreceptor cells, revealed by expression levels of marker proteins and by retinal cell proliferation.
There are some gaps in our knowledge about how TH levels affect the eyes of already fully developed organisms and/or whether they have similarly serious effects on the retinal layers. It can be assumed that the effects, if any, are weaker.
Also, the molecular process of how a lower TH level leads to disturbances of the layers in the eye is not yet fully understood.
There is little mammalian data on this KER.
Also, studies (Reider et al. 2014) found that layer thickness varied across the ages suggesting that these retinal layers are differentially sensitive to for example MMI and/or that there are different critical periods of sensitiv ity in retinal tissue.
Quantitative Understanding of the Linkage
Known Feedforward/Feedback loops influencing this KER- One feedback loop mechanism could be triggered by iodine deficiency or inhibition of iodine uptake. It appears probably that the inhibition increases the secretion of Thyroid stimulating hormone, which could stimulate the expression of the NIS-transporter. This increase in TSH could shift the ratio in favour of T3.
References
Baumann L, Ros A, Rehberger K, Neuhauss SCF, Segner H. 2016. Thyroid disruption in zebrafish (Danio rerio) larvae: Different molecular response patterns lead to impaired eye development and visual functions. Aquatic Toxicology 172:44-55.
Besson, M., Feeney, W. E., Moniz, I., François, L., Brooker, R. M., Holzer, G., Metian, M., Roux, N., Laudet, V., & Lecchini, D. (2020). Anthropogenic stressors impact fish sensory development and survival via thyroid disruption. Nature Communications, 11(1). https://doi.org/10.1038/s41467-020-17450-8
Bhumika, S., & Darras, V. M. (2014). Role of thyroid hormones in different aspects of nervous system regeneration in vertebrates. General and Comparative Endocrinology, 203, 86–94. https://doi.org/10.1016/j.ygcen.2014.03.017
Duval, M. G., & Allison, W. T. (2018). Photoreceptor progenitors depend upon coordination of gdf6a, thrβ, and tbx2b to generate precise populations of cone photoreceptor subtypes. Investigative Ophthalmology and Visual Science, 59(15), 6089–6101. https://doi.org/10.1167/iovs.18-24461
Darras VM, Houbrechts AM, Van Herck SLJ. Intracellular thyroid hormone metabolism as a local regulator of nuclear thyroid hormone receptor-mediated impact on vertebrate development. Biochim Biophys Acta - Gene Regul Mech. 2015;1849(2):130-141. doi:10.1016/j.bbagrm.2014.05.004
Gan, K. J., & Flamarique, I. N. (2010). Thyroid hormone accelerates opsin expression during early photoreceptor differentiation and induces opsin switching in differentiated TRα-expressing cones of the salmonid retina. Developmental Dynamics, 239(10), 2700–2713. https://doi.org/10.1002/dvdy.22392
Houbrechts AM, Delarue J, Gabriëls IJ, Sourbron J, Darras VM. Permanent deiodinase type 2 Deficiency strongly perturbs zebrafish development, growth, and fertility. Endocrinology. 2016;157(9):3668-3681. doi:10.1210/en.2016-1077
Komoike Y, Matsuoka M, Kosaki K. 2013. Potential Teratogenicity of Methimazole: Exposure of Zebrafish Embryos to Methimazole Causes Similar Developmental Anomalies to Human Methimazole Embryopathy. Birth Defects Research Part B-Developmental and Reproductive Toxicology 98(3):222-229.
Gamborino MJ, Sevilla-Romero E, Muñoz A, Hernández-Yago J, Renau-Piqueras J, Pinazo-Durán MD. Role of thyroid hormone in craniofacial and eye development using a rat model. Ophthalmic Res. 2001;33(5):283-291. doi:10.1159/000055682
Reider M, Connaughton VP. 2014. Effects of Low-Dose Embryonic Thyroid Disruption and Rearing Temperature on the Development of the Eye and Retina in Zebrafish. Birth Defects Research Part B-Developmental and Reproductive Toxicology 101(5):347-354.
Quesada-García A, Encinas P, Valdehita A, et al. Thyroid active agents T3 and PTU differentially affect immune gene transcripts in the head kidney of rainbow trout (Oncorynchus mykiss). Aquat Toxicol. 2016;174:159-168. doi:10.1016/j.aquatox.2016.02.016
Wang, W. Der, Hsu, H. J., Li, Y. F., & Wu, C. Y. (2017). Retinoic acid protects and rescues the development of zebrafish embryonic retinal photoreceptor cells from exposure to paclobutrazol. International Journal of Molecular Sciences, 18(1). https://doi.org/10.3390/ijms18010130
Relationship: 2374: Altered, retinal layer structure leads to Altered, Visual function
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Thyroperoxidase inhibition leading to altered visual function via altered retinal layer structure | adjacent |
Evidence Supporting Applicability of this Relationship
| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| zebrafish | Danio rerio | NCBI |
| Life Stage | Evidence |
|---|---|
| Embryo | High |
| Adult | High |
| Juvenile |
| Sex | Evidence |
|---|---|
| Unspecific |
Taxonomic applicability: The visual system of the zebrafish follows the typical organisation of vertebrates and is often used as a model to study human eye diseases. Although there are some differences in eye structure between zebrafish and humans, it is plausible to assume that a functioning eye structure is important for visual function across all vertebrates and invertebrates that have eyes.
Life stage applicability: The first visual responses based on retinal functionality appear around 70 hpf in zebrafish (Schmitt and Dowling 1999). It is plausible to assume that alterations of the eye structure would result in altered visual function across all life stages, but such alterations are most likely to occur during the development of the normal eye structure, which occurs in the embryo-eleutheroembryo phase.
Sex applicability: Zebrafish are undifferentiated gonochorists since both sexes initially develop an immature ovary (Maack and Segner, 2003). Immature ovary development progresses until approximately the onset of the third week. Later, in female fish immature ovaries continue to develop further, while male fish undergo transformation of ovaries into testes. Final transformation into testes varies among male individuals, however finishes usually around 6 weeks post fertilization. Effects on visual function resulting from altered eye structure during early development are therefore expected to be independent of sex.
Key Event Relationship Description
The structure of the vertebrate retina is well conserved and consists of the following layers: The retinal pigment epithelium (RPE), the photoreceptor layer (PRL), the outer plexiform layer (OPL), the inner nuclear layer (INL), the inner plexiform layer (IPL) and the ganglion cell layer (GCL). Each of these layers has a specific function for the physiology of the visual system. The RPE serves to protect and maintain the photoreceptors and absorbs excess light. The photoreceptors in the PRL consist of a light-receiving outer segment (OS) and the inner segment (IS), which contains the cell bodies. They send their signals to the bipolar cells in the INL, which transmit the signal to the ganglion cells. These form the optic nerve and are responsible for transmitting signals to the optic nerves. In both plexiform layers, the retinal neurons form their synaptic connections (Bibliowicz et al. 2011).
To study the eye, the zebrafish (Danio rerio) is at the forefront of many studies as a model organism. In zebrafish, eye development begins around 12 hpf (Houbrechts et al., 2016b) and by 72 hpf the layers of the retina are well developed (Malicki et al., 2016). Functional vision is established by 4-5 dpf (Brockerhoff, 2006; Chhetri et al., 2014).
Evidence Supporting this KER
Biological PlausibilityIt should be emphasised that all layers of the retina are interdependent. The RPE plays an essential role in the retinoid cycle for the photoreceptors (PRL), which perceive the light stimulus and transmit it via the bipolar cells to the ganglia (IPL), which form the optic nerve and transmit the signal to the optic nerve (Connaughton 2005). If these key sites of the phototransduction pathway are disrupted by, for example, endocrine disruptors, it stands to reason that there would be a significant impact on the optical sense and it is plausible that disorders of the eye structure can lead to visual disorders.
-
Avallone et al. (2015) studied the effects of cadmium exposure on the vision of adult zebrafish. The morpho-cytological changes of the retina (Nerve fiber layer clearly thickened and vacuolated, presence of compact pycnotic nuclei, empty area, change in the thickness of pigmented retinal epithelium and at the level of cones inner segments, extended folds of threated retinae, presence of cell debris and/of blood cells in vitreal chamber) were investigated by light and electron microscopy, while the functionality of the cadmium-exposed retinas was assessed by re-illumination behavioural tests with white or coloured light. Cadmium toxicity was shown to cause significant cell degeneration and loss of organisation at both macroscopic and microscopic levels. These changes were directly related to functional responses, particularly by increasing light sensitivity of exposed fish.
-
Baumann et al., 2016 used propylthiouracil (PTU) and tetrabromobisphenol A (TBBPA) to disrupt the thyroid hormone system in zebrafish larvae. This exposure induced different molecular response patterns leading to impaired eye development (reduction of RPE cell diameter, pigmentation and eye size). Behavioural analyses showed that these larvae were also disrupted in their visual capacities, such as decrease in optokinetic response and increase in light preference of PTU-treated larvae.
-
Chawla et al., 2018 showed that Retinoic Acid (RA) maintains function of neural crest-derived ocular and craniofacial structures in adult zebrafish. An increase in RA caused morphological changes: prognathic jaw and increased distance from the eyes to the gill. The inhibition of RA increased the head size. This change of dimensions of craniofacial structures and the change of RA levels showed decrease aqueous outflow from anterior segment of the eye together with the decrease of visual function (Optokinetic reflex test).
-
Crowley-Perry et al. (2021) showed that transient, developmental exposure to sublethal and environmentally relevant concentrations of bisphenol A (BPA) alters larval eye morphology and visually guided behaviors. BPA exposure increased eye diameters and the number of larvae displaying a positive optomotor response (OMR).
-
Flamarique et al. (2013b) used thyroid hormone treatment to transform the UV cones of young rainbow trout into blue cones and showed that this reduced the distances and angles at which prey were located (variables that are known indicators of foraging performance). Using optical measurements and photon-catch calculations, the study showed that control rainbow trouts perceived prey (Daphnia) with greater contrast compared to thyroid-hormone-treated fish, demonstrating that the presence of UV cones enhances foraging performance of young rainbow trout.
-
Heijlen et al. (2014) showed that knockdown of Type 3 Iodothyronine Deiodinase severely perturbs both embryonic and early larval development in zebrafish. Furthermore, D3-knockdown larvae spent significantly less time moving, and both embryos and larvae exhibited perturbed escape responses, suggesting that D3 knockdown affects muscle development and/or functioning.
-
Houbrechts et al. (2016) used a knockdown of deiodinase 1 and 2 genes in zebrafish embryos to induce transient hypothyroidism and observed decreased levels of mRNA coding for rod and cone opsins (at 3 dpf, days post fertilization) and a strong transient reduction in rods and all four cone types (at 3 dpf but no longer at 7 dpf) together with a transiently reduced response (increase of swimming activity) to light (4 dpf, but no longer at 7 dpf).
-
Masuda et al. (2017) and Naujokas et al. (2013) found out that retinal diseases could eventually lead to vision loss and blindness, with signifying symptoms of inflammation and oxidative stress in retinal cells or tissue
-
Van Camp et al. (2018) showed that permanent deiodinase 2 deficiency in zebrafish resulted in a reduction of the number of R/G cones and rods that persisted through 7 dpf together with a reduced response to light (observed at 6 dpf).
-
Walter et al. (2019) found out that developmental exposure to either T4 or T3 in zebrafish embyros altered photomotor behavior in a concentration- and time-dependent manner. T4 increased locomotor activity in response to sudden changes from light to dark. T3 caused similar patterns of changes in photomotor behavior at 3 and 4 dpf.
-
Frau et al. (2020) studied the consequences of differences in photoreceptor patterning across fish species. They concluded that species that are primarily nocturnal or live in low light environments such as the common sole and Senegalese sole have a less ordered mosaic cone pattern. A study of different fish species revealed that lattice-like patterning of the cone mosaic seems to improve visual acuity. Fish taxa that live in low light environments generally do not possess lattice-like cone mosaics.
Often, high variances occur in the results of behavioural studies because an organism has some compensatory mechanisms that allow it to survive.
It is more difficult to compare data from different laboratories in such experiments.
Similarly, extrapolating data from fish to mammalian data is particularly difficult for behavioural studies.
References
Strauss O. The retinal pigment epithelium in visual function. Physiol Rev. 2005 Jul;85(3):845-81. doi: 10.1152/physrev.00021.2004. PMID: 15987797.
Avallone B, Crispino R, Cerciello R, Simoniello P, Panzuto R, Motta CM. Cadmium effects on the retina of adult Danio rerio. CR Biologies. 2015;338:40–7.
Baumann, L., Ros, A., Rehberger, K., Neuhauss, S. C. F., & Segner, H. (2016). Thyroid disruption in zebrafish (Danio rerio) larvae: Different molecular response patterns lead to impaired eye development and visual functions. Aquatic Toxicology, 172, 44–55. https://doi.org/10.1016/j.aquatox.2015.12.015
Bibliowicz, J., Tittle, R. K., & Gross, J. M. (2011). Toward a better understanding of human eye disease: Insights from the zebrafish, Danio rerio. In Progress in Molecular Biology and Translational Science(Vol. 100, Issue Table 1). https://doi.org/10.1016/B978-0-12-384878-9.00007-8
Brockerhoff, S. E. (2006). Measuring the optokinetic response of zebrafish larvae. Nature Protocols, 1(5), 2448–2451. https://doi.org/10.1038/nprot.2006.255
Chawla, B., Swain, W., Williams, A. L., & Bohnsack, B. L. (2018). Retinoic acid maintains function of neural crest–derived ocular and craniofacial structures in adult zebrafish. Investigative Ophthalmology and Visual Science, 59(5), 1924–1935. https://doi.org/10.1167/iovs.17-22845
Chhetri, J., Jacobson, G., & Gueven, N. (2014). Zebrafish-on the move towards ophthalmological research. Eye (Basingstoke), 28(4), 367–380. https://doi.org/10.1038/eye.2014.19
Crowley-Perry, M., Barberio, A. J., Zeino, J., Winston, E. R., & Connaughton, V. P. (2021). Zebrafish optomotor response and morphology are altered by transient, developmental exposure to bisphenol-a. Journal of Developmental Biology,9(2). https://doi.org/10.3390/jdb9020014
Connaughton, V. P., Graham, D., & Nelson, R. (2004). Identification and morphological classification of horizontal, bipolar, and amacrine cells within the zebrafish retina. Journal of Comparative Neurology, 477(4), 371–385. https://doi.org/10.1002/cne.20261
Flamarique, I. N. (2013). Opsin switch reveals function of the ultraviolet cone in fish foraging. Proceedings of the Royal Society B: Biological Sciences, 280(1752). https://doi.org/10.1098/rspb.2012.2490
Frau, S., Novales Flamarique, I., Keeley, P. W., Reese, B. E., & Muñoz-Cueto, J. A. (2020). Straying from the flatfish retinal plan: Cone photoreceptor patterning in the common sole (Solea solea) and the Senegalese sole (Solea senegalensis). Journal of Comparative Neurology, 528(14), 2283–2307. https://doi.org/10.1002/cne.24893
Heijlen, M., Houbrechts, A. M., Bagci, E., Van Herck, S. L. J., Kersseboom, S., Esguerra, C. V., Blust, R., Visser, T. J., Knapen, D., & Darras, V. M. (2014). Knockdown of type 3 iodothyronine deiodinase severely perturbs both embryonic and early larval developme
Houbrechts, A. M., Vergauwen, L., Bagci, E., Van houcke, J., Heijlen, M., Kulemeka, B., Hyde, D. R., Knapen, D., & Darras, V. M. (2016). Deiodinase knockdown affects zebrafish eye development at the level of gene expression, morphology and function. Molecular and Cellular Endocrinology, 424, 81–93. https://doi.org/10.1016/j.mce.2016.01.018
Malicki, J., Pooranachandran, N., Nikolaev, A., Fang, X., & Avanesov, A. (2016). Analysis of the retina in the zebrafish model. In Methods in Cell Biology(Vol. 134). Elsevier Ltd. https://doi.org/10.1016/bs.mcb.2016.04.017
Masuda, T., Shimazawa, M., Hara, H., 2017. Retinal diseases associated with oxidative stress and the effects of a free radical scavenger (edaravone). Oxid. Med. Cell. Longev. 2017, 9208489.
Moreno-Marmol T, Cavodeassi F, Bovolenta P. 2018. Setting Eyes on the Retinal Pigment Epithelium. Frontiers in Cell and Developmental Biology 6.
Morris AC, Fadool JM. 2005. Studying rod photoreceptor development in zebrafish. Physiology & Behavior 86(3):306-313.
Naujokas, M.F., Anderson, B., Ahsan, H., et al., 2013. The broad scope of health effects from chronic arsenic exposure: update on a worldwide public health problem. Environ. Health Perspect. 121 (3), 295e302.
Raymond PA, Barthel LK, Curran GA. 1995. DEVELOPMENTAL PATTERNING OF ROD AND CONE PHOTORECEPTORS IN EMBRYONIC ZEBRAFISH. Journal of Comparative Neurology 359(4):537-550.
Schmitt EA, Dowling JE. 1999. Early retinal development in the zebrafish, Danio rerio: Light and electron microscopic analyses. Journal of Comparative Neurology 404(4):515-536.
Vancamp P, Houbrechts AM, Darras VM. Insights from zebrafish deficiency models to understand the impact of local thyroid hormone regulator action on early development. Gen Comp Endocrinol. 2019 Aug 1;279:45-52. doi: 10.1016/j.ygcen.2018.09.011. Epub 2018 Sep 19. PMID: 30244055.
Walter, K. M., Miller, G. W., Chen, X., Harvey, D. J., Puschner, B., & Lein, P. J. (2019). Changes in thyroid hormone activity disrupt photomotor behavior of larval zebrafish. Neurotoxicology, 74, 47–57. https://doi.org/10.1016/j.neuro.2019.05.008
Relationship: 2375: Altered, Visual function leads to Increased Mortality
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Inhibition of Fyna leading to increased mortality via decreased eye size (Microphthalmos) | adjacent | ||
| Thyroperoxidase inhibition leading to altered visual function via altered retinal layer structure | adjacent | ||
| Thyroperoxidase inhibition leading to altered visual function via decreased eye size | adjacent | ||
| Thyroperoxidase inhibition leading to altered visual function via altered photoreceptor patterning | adjacent |
Evidence Supporting Applicability of this Relationship
| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| zebrafish | Danio rerio | NCBI |
| Life Stage | Evidence |
|---|---|
| All life stages |
| Sex | Evidence |
|---|---|
| Unspecific |
Taxonomic applicability: The visual system of the fish (e.g., zebrafish) follows the typical organisation of vertebrates and is often used as a model to study human eye diseases. Although there are some differences, it is plausible to assume that visual function is important for survival across all vertebrates and invertebrates that have eyes.
Sex applicability: Zebrafish are undifferentiated gonochorists since both sexes initially develop an immature ovary (Maack and Segner, 2003). Immature ovary development progresses until approximately the onset of the third week. Later, in female fish immature ovaries continue to develop further, while male fish undergo transformation of ovaries into testes. Final transformation into testes varies among male individuals, however finishes usually around 6 weeks post fertilization. Effects on mortality resulting from altered visual function are therefore expected to be independent of sex.
Life stage applicability: It is plausible to assume that altered visual function of the eye would result in a higher mortality across all life stages. This could be especially true for the embryonic stages, the most sensitive stage of life. Vision plays a crucial role (in the early life stages) of most species, as eye development and establishment of functional vision is essential for perception of food or avoidance of predators for example (Carvalho et al., 2002).
Key Event Relationship Description
In animals, whatever the taxa, visual abilities are strongly linked to their lifestyle (feeding, avoidance of predators, movement, protection....). When these capacities are impaired, they lead to reduced fitness and are therefore strongly linked to a decrease in survival, particularly in the early stages of life.
Evidence Supporting this KER
Biological PlausibilityDecreases in visual functions can have a strong impact on behavior, leading to changes in individual response and abilities in the environment, including, for example, perception of food or avoidance of predators. Variation in the visual system can also influence learning tasks when visual stimuli are used (Corral-López et al., 2017).
Sensory drive has been implicated in speciation in various taxa, largely based on phenotype-environment correlations and signatures of selection in sensory genes, including view (Maan et al, 2017).
It can be assumed that an animal which has difficulties in finding food and avoiding predators will have lower survival chances in wildlife.
Only very few studies are available in which it was demonstrated that decreased visual capacities lead to reduced survival of the organism. In general, mortality is rarely assessed but survival-reducing factors (feeding, predation) are mainly investigated. Here we consider the work about different toxicants that disrupt complex fish behaviors, such as predator avoidance, reproductive, and social behaviors. Toxicant exposure often completely eliminates the performance of behaviors that are essential to fitness and survival in natural ecosystems, frequently after exposures of lesser magnitude than those causing significant mortality (Brown et al., 2004).
-
(Dehnert et al., 2019): In zebrafish, 2, 4-Dichlorophenoxyacetic exposure during eye development impaired visual behavior, i.e. reduced prey capture. Additonally, exposed fish showed reduced neural activity within the optic tectum following prey exposure.
-
(Besson et al., 2020) exposed metamorphosing convict surgeonfish to either chlorpyrifos or elevated levels of T3. Sensory organ development (eyes, lateral line, olfactory epithelium) was severely impaired and fish displayed reduced reaction to artificial predator cues. A predation test in an in situ arena revealed that T3-treated fish exhibited significantly higher survival than control fish, while chlorpyriphos-treated fish showed reduced survival.
-
(Babkiewicz et al., 2020) investigated the effect of temperature on the visual detection distance of zebrafish (Danio rerio) larvae by measuring the largest distance from a moving target that induced a neural response in the optic tectum. Fish raised in elevated temperatures showed reduced visual detection distance.
It is obvious that impaired vision leads to higher mortality, as the sense of sight is important for survival, and if it is impaired, feeding or escape becomes more difficult. However, studies investigating this connection are still very rare. Also, the natural conditions, which depend on many variables, are difficult to reproduce in the laboratory or to compare within different laboratories.
Quantitative Understanding of the Linkage
Known modulating factorsIncrease according to global health of the population (e.g on trout (Post and Parkinson, 2001)
References
Babkiewicz, E., Bazała, M., Urban, P., Maszczyk, P., Markowska, M., & Maciej Gliwicz, Z. (2020). The effects of temperature on the proxies of visual detection of Danio rerio larvae: observations from the optic tectum. Biology Open, 9(7). https://doi.org/10.1242/BIO.047779
Besson, M., Feeney, W. E., Moniz, I., François, L., Brooker, R. M., Holzer, G., Metian, M., Roux, N., Laudet, V., & Lecchini, D. (2020). Anthropogenic stressors impact fish sensory development and survival via thyroid disruption. Nature Communications, 11(1). https://doi.org/10.1038/s41467-020-17450-8
Brown, S. B., Adams, B. A., Cyr, D. G., & Eales, J. G. (2004). Contaminant effects on the teleost fish thyroid. Environmental Toxicology and Chemistry, 23(7), 1680–1701. https://doi.org/10.1897/03-242
Carvalho, P. S. M., Noltie, D. B., & Tillitt, D. E. (2002). Ontogenetic improvement of visual function in the medaka Oryzias latipes based on an optomotor testing system for larval and adult fish. Animal Behaviour, 64(1), 1–10. https://doi.org/10.1006/anbe.2002.3028
Corral-López, A., Garate-Olaizola, M., Buechel, S. D., Kolm, N., & Kotrschal, A. (2017). On the role of body size, brain size, and eye size in visual acuity. Behavioral Ecology and Sociobiology, 71(12). https://doi.org/10.1007/s00265-017-2408-z
Dehnert, G. K., Karasov, W. H., & Wolman, M. A. (2019). 2,4-Dichlorophenoxyacetic acid containing herbicide impairs essential visually guided behaviors of larval fish. Aquatic Toxicology, 209(October 2018), 1–12. https://doi.org/10.1016/j.aquatox.2019.01.015
Relationship: 2013: Increased Mortality leads to Decrease, Population trajectory
AOPs Referencing Relationship
Evidence Supporting Applicability of this Relationship
| Life Stage | Evidence |
|---|---|
| All life stages | High |
| Sex | Evidence |
|---|---|
| Unspecific | Moderate |
Taxonomic: All organisms must survive to reproductive age in order to reproduce and sustain populations. The additional considerations related to survival made above are applicable to other fish species in addition to zebrafish and fathead minnows with the same reproductive strategy (r-strategist as described in the theory of MaxArthur and Wilson (1967). The impact of reduced survival on population size is even greater for k-strategists that invest more energy in a lower number of offspring.
Life stage: Density dependent effects start to play a role in the larval stage of fish when free-feeding starts (Hazlerigg et al., 2014).
Sex: This linkage is independent of sex.
Key Event Relationship Description
Increased mortality in the reproductive population may lead to a declining population. This depends on the excess mortality due to the applied stressor and the environmental parameters such as food availability and predation rate. Most fish species are r-strategist, meaning they produce a lot of offspring instead of investing in parental care. This results in natural high larval mortality causing only a small percentage of the larvae to survive to maturity. If the excess larval mortality due to a stressor is small, the population dynamics might result in constant population size. Should the larval excess be more significant, or last on the long-term, this will affect the population. To calculate the long-term persistence of the population, population dynamic models should be used.
Evidence Supporting this KER
Survival rate is an obvious determinant of population size and is therefore included in population modeling (e.g., Miller et al., 2020).
Biological Plausibility- Survival to reproductive maturity is a parameter of demographic significance. Assuming resource availability (i.e., food, habitat, etc.) is not limiting to the extant population, sufficient mortality in the reproductive population may ultimately lead to declining population trajectories.
- Under some conditions, reduced larval survival may be compensated by reduced predation and increased food availability, and therefore not result in population decline (Stige et al., 2019).
- According to empirical data, combined with population dynamic models, feeding larvae are the crucial life stage in zebrafish (and other r-strategists) for the regulation of the population. (Schäfers et al., 1993)
- In fathead minnow, natural survival of early life stages has been found to be highly variable and influential on population growth (Miller and Ankley, 2004)
- Rearick et al. (2018) used linked data from behavioural assays to survival trials and applied a modelling approach to quantify changes in antipredator escape performance of larval fathead minnows in order to predict changes in population abundance. This work was done in the context of exposure to an environmental oestrogen. Expsoed fish had delayed response times and slower escape speeds, and were more susceptible to predation. Population modelling showed that his can result in population decline.
- In the context of fishing and fisheries, ample evidence of a link between increased mortality and a decrerase of population size has been given. Important insights can result from the investigation of optimum modes of fishing that allow for maintaining a population (Alekseeva and Rudenko, 2018). Jacobsen and Essington (2018) showed the impact of varying predation mortality on forage fish populations.
- Boreman (1997) reviewed methods for comparing the population-level effects of mortality in fish populations induced by pollution or fishing.
-
The extent to which larval mortality affects population size could depend on the fraction of surplus mortality compared to a natural situation.
-
There are scenarios in which individual mortality may not lead to declining population size. These include instances where populations are limited by the availability of habitat and food resources, which can be replenished through immigration. Effects of mortality in the larvae can be compensated by reduced competition for resources (Stige et al., 2019).
-
The direct impact of pesticides on migration behavior can be difficult to track in the field, and documentation of mortality during migration is likely underestimated (Eng 2017).
References
Alekseeva SM, Rudenko AI. 2018. Modeling of optimum fishing population. Marine Intellectual Technologies. 3(4):142-146.
Beaudouin, R., Goussen, B., Piccini, B., Augustine, S., Devillers, J., Brion, F., Pery, A.R., 2015. An individual-based model of zebrafish population dynamics accounting for energy dynamics. PloS one 10, e0125841.
Boreman J. 1997. Methods for comparing the impacts of pollution and fishing on fish populations. Transactions of the American Fisheries Society. 126(3):506-513.
Caswell, H., 2000. Matrix population models. Sinauer Sunderland, MA, USA.
Eng, M.L., Stutchbury, B.J.M. & Morrissey, C.A. Imidacloprid and chlorpyrifos insecticides impair migratory ability in a seed-eating songbird. Sci Rep 7, 15176 (2017)
Hazlerigg, C.R., Lorenzen, K., Thorbek, P., Wheeler, J.R., Tyler, C.R., 2012. Density-dependent processes in the life history of fishes: evidence from laboratory populations of zebrafish Danio rerio. PLoS One 7, e37550.
Jacobsen NS, Essington TE. 2018. Natural mortality augments population fluctuations of forage fish. Fish and Fisheries. 19(5):791-797.
MacArthur, R., Wilson, E., 1967. The Theory of Island Biogeography. Princeton: Princeton Univ. Press. 203 p.
Miller, D.H., Ankley, G.T., 2004. Modeling impacts on populations: fathead minnow (Pimephales promelas) exposure to the endocrine disruptor 17β-trenbolone as a case study. Ecotoxicology and Environmental Safety 59, 1-9.
Miller, D.H., Clark, B.W., Nacci, D.E. 2020. A multidimensional density dependent matrix population model for assessing risk of stressors to fish populations. Ecotoxicology and environmental safety 201, 110786
Pinceel, T., Vanschoenwinkel, B., Brendonck, L., Buschke, F., 2016. Modelling the sensitivity of life history traits to climate change in a temporary pool crustacean. Scientific reports 6, 29451.
Rearick, D.C., Ward, J., Venturelli, P., Schoenfuss, H., 2018. Environmental oestrogens cause predation-induced population decline in a freshwater fish. Royal Society open science 5, 181065.
Schäfers, C., Oertel, D., Nagel, R., 1993. Effects of 3, 4-dichloroaniline on fish populations with differing strategies of reproduction. Ecotoxicology and Ecophysiology, 133-146.
Stige, L.C., Rogers, L.A., Neuheimer, A.B., Hunsicker, M.E., Yaragina, N.A., Ottersen, G., Ciannelli, L., Langangen, Ø., Durant, J.M., 2019. Density‐and size‐dependent mortality in fish early life stages. Fish and Fisheries 20, 962-976.Hazlerigg, C.R.E., Tyler, C.R., Lorenzen, K., Wheeler, J.R., Thorbek, P., 2014. Population relevance of toxicant mediated changes in sex ratio in fish: An assessment using an individual-based zebrafish (Danio rerio) model. Ecological Modelling 280, 76-88.
Stige, L.C., Rogers, L.A., Neuheimer, A.B., Hunsicker, M.E., Yaragina, N.A., Ottersen, G., Ciannelli, L., Langangen, O., Durant, J.M., 2019. Density- and size-dependent mortality in fish early life stages. Fish and Fisheries 20, 962-976.
List of Non Adjacent Key Event Relationships
Relationship: 366: Thyroperoxidase, Inhibition leads to T4 in serum, Decreased
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Inhibition of Thyroperoxidase and Subsequent Adverse Neurodevelopmental Outcomes in Mammals | non-adjacent | High | Moderate |
| Thyroperoxidase inhibition leading to increased mortality via reduced anterior swim bladder inflation | non-adjacent | High | Low |
| Thyroperoxidase inhibition leading to altered amphibian metamorphosis | non-adjacent | High | High |
| Thyroperoxidase inhibition leading to altered visual function via altered retinal layer structure | non-adjacent | High | Low |
Evidence Supporting Applicability of this Relationship
| Life Stage | Evidence |
|---|---|
| All life stages | High |
| Sex | Evidence |
|---|---|
| Male | High |
| Female | High |
Taxonomic: Use of TPO inhibitors as anti-hyperthyroidism drugs in humans and pets (Emiliano et al., 2010; Trepanier, 2006) and effects of these drugs on serum TH concentrations in rats (US EPA, 2005), amphibian, fish and avian species (Coady et al., 2010; Grommen et al., 2011; Nelson et al., 2016; Rosebrough et al., 2006; Stinckens et al.; 2020; Tietge et al., 2012), strongly supports a causative linkage between inhibition of TPO and decreased serum T4 across species. Therefore, this KER is plausibly applicable across vertebrate species. Therefore, this KER is plausibly applicable across vertebrates.
Life stage: Applicability to certain life stages may depend on the species and their dependence on maternally transferred thyroid hormones during the earliest phases of development. The earliest life stages of teleost fish rely on maternally transferred THs to regulate certain developmental processes until embryonic TH synthesis is active (Power et al., 2001). As a result, TPO inhibition is not expected to decrease TH synthesis during these earliest stages of development. In zebrafish, Opitz et al. (2011) showed the formation of a first thyroid follicle at 55 hours post fertilization (hpf), Chang et al. (2012) showed a first significant TH increase at 120 hpf and Walter et al. (2019) showed clear TH production already at 72 hpf but did not analyse time points between 24 and 72 hpf. In fathead minnows, a significant increase of whole body thyroid hormone levels was already observed between 1 and 2 dpf, which corresponds to the appearance of the thyroid anlage at 35 hpf prior to the first observation of thyroid follicles at 58 hpf (Wabuke-Bunoti and Firling, 1983). Therefore, iIt is still uncertain when exactly embryonic TH synthesis is activated and how this determines sensitivity to TH disruptors.
Sex: The KE is plausibly applicable to both sexes. Thyroid hormones are essential in both sexes and the components of the HPT-axis are identical in both sexes. There can however be sex-dependent differences in the sensitivity to the disruption of thyroid hormone levels and the magnitude of the response. In humans, females appear more susceptible to hypothyroidism compared to males when exposed to certain halogenated chemicals (Hernandez‐Mariano et al., 2017; Webster et al., 2014). In adult zebrafish, Liu et al. (2019) showed sex-dependent changes in thyroid hormone levels and mRNA expression of regulatory genes including corticotropin releasing hormone (crh), thyroid stimulating hormone (tsh) and deiodinase 2 after exposure to organophosphate flame retardants. The underlying mechanism of any sex-related differences remains unclear.
Key Event Relationship Description
Thyroperoxidase (TPO) is the enzyme that catalyzes iodine organification of thyroglobulin to produce thyroglobulin (Tg)-bound T3 and T4 in the lumen of thyroid follicles. Tg-bound THs are endocytosed across the apical lumen-follicular cell membrane, undergo thyroglobulin proteolysis, followed by hormone section into the blood stream (see Taurog, 2005 for review). This indirect KER describes the relationship of TPO inhibition to reduced circulating levels of thyroid hormone (TH) in the serum.
Evidence Supporting this KER
The weight of evidence linking thyroperoxidase inhibition to reductions in circulating serum TH is strong. Many studies support this basic linkage. There is no inconsistent data.
Biological PlausibilityIt is a well-accepted fact that inhibition of the only enzyme capable of synthesizing THs, TPO, results in subsequent decrease in serum TH concentrations. A large amount of evidence from clinical and animal studies clearly support the commonly accepted dogma that inhibition of TPO leads to decreased serum THs.
Empirical EvidenceThe majority of research in support of this KER involve exposure to known TPO inhibitors and measurement of serum hormones. There are many in vivo studies that link decreases in serum TH concentrations with exposure to xenobiotics that inhibit thyroperoxidase (TPO) in mammals (Brucker-Davis, 1998; Hurley, 1998; Boas et al., 2006; Crofton, 2008; Kohrle, 2008; Pearce and Braverman, 2009; Murk et al., 2013).
While these studies support the connection between exposure to a known TPO inhibitor and decreased TH, many of these studies do not empirically measure TPO inhibition or decreased TH synthesis. Thus, many studies support the indirect linkage between TPO inhibition (for chemicals identified as TPO inhibitors in in vivo or ex vivo studies) and decreased TH, with the well accepted theory that these proceed via decreased TH synthesis. That exposure to TPO inhibitors leads to decreased serum TH concentrations, via decreased TH synthesis is strongly supported by decades of mechanistic research in a variety of species.
This indirect relationship is also evidenced by the use of clinically-relevant anti-hyperthyroidism drugs, MMI and PTU (Laurberg & Anderson, 2014; Sundaresh et al., 2013). These drugs are both recognized TPO inhibitors and are part of a standard drug-based regimen of care for clinically hyperthyroid patients including those with Grave's disease. Serum THs are measured as the bioindicator of successful treatment with anti-hyperthyroidism drugs; the actual decrease in TH synthesis in the thyroid gland is implied in the efficacious use of these drugs (Trepanier, 2006).
In rats, MMI and PTU are often used as control chemicals to decrease serum THs to study biological phenomena related to disruption of TH homeostasis (many examples, including Zoeller and Crofton, 2005; Morreale de Escobar et al, 2004; Schwartz et al., 1997; Herwig et al., 2014; Wu et al., 2013; Pathak et al., 2011). Further, MMI is recommended as a positive control for use in the Amphibian Metamorphosis (Frog) Assay within Tier 1 of the U.S. EPA Endocrine Disruptor Screening Program (US EPA, 2009; Coady et al., 2010), an assay used to evaluate the potential for chemicals to disrupt TH homeostasis. PTU has been suggested as positive control chemical in the guidance for the Comparative Developmental Thyroid Assay (US EPA, 2005), a non-guideline assay used to evaluate the potential for chemicals to disrupt TH homeostasis during gestation and early neonatal development.
Additionally, evidence is available from studies investigating responses to TPO inhibitors in fish. For example, Stinckens et al. (2020) showed reduced whole body T4 concentrations in zebrafish larvae exposed to 50 or 100 mg/L methimazole, a potent TPO inhibitor, from immediately after fertilization until 21 or 32 days of age. Exposure to 37 or 111 mg/L propylthiouracil also reduced T4 levels after exposure up to 14, 21 and 32 days in the same study. Walter et al. (2019) showed that propylthiouracil had no effect on T4 levels in 24h old zebrafish, but decreased T4 levels of 72h old zebrafish. This difference is probably due to the onset of embryonic TH production between the age of 24 and 72 hours (Opitz et al., 2011). Stinckens et al. (2016) showed that exposure to 2-mercaptobenzothiazole (MBT), an environmentally relevant TPO inhibitor, decreased whole body T4 levels in continuously exposed 5 and 32 day old zebrafish larvae. A high concentration of MBT also decreased whole body T4 levels in 6 day old fathead minnows, but recovery was observed at the age of 21 days although the fish were kept in the exposure medium (Nelson et al., 2016). Crane et al. (2006) showed decreased T4 levels in 28 day old fathead minnows continuously exposed to 32 or 100 µg/L methimazole.
Thus, an indirect key event relationship between TPO inhibition and decreased serum THs is strongly supported by a large database of clinical medicine and investigative research with whole animals (with a great deal of supporting evidence in rats and frogs).
Temporal Evidence: In mammals, the temporal nature of this KER is applicable to all life stages, including development (Seed et al., 2005). The qualitative impact of thyroperoxidase inhibition on serum hormones is similar across all ages in mammals. The temporal nature of the impact on serum THs by TPO inhibitors in developmental exposure studies is evidenced by the duration of exposure and developmental age (Goldey et al., 1995; Ahmed et al., 2010; Tietge et al., 2010), as well as recovery after cessation of exposure (Cooke et al., 1993; Goldey et al., 1995; Sawin et al., 1998; Axelstad et al., 2008; Shibutani et al., 2009; Lasley and Gilbert, 2011). The temporal relationship between TPO inhibitor exposure duration and serum hormone decreases in adult organisms has been widely demonstrated (e.g., Hood et al., 1999; Mannisto et al., 1979). In addition, MMI and PTU induced decreases in serum T4 are alleviated by thyroid hormone replacement in both fetal and postnatal age rats (Calvo et al., 1990; Sack et al., 1995; Goldey and Crofton, 1998). Computational modeling of the thyroid also provides evidence for the indirect temporal relationship between these two KEs (e.g., Degon et al., 2008; Fisher et a., 2013).
In oviparous fish such as zebrafish and fathead minnow, the nature of this KER depends on the life stage since the earliest stages of embryonic development rely on maternal thyroid hormones transferred to the eggs. Embryonic thyroid hormone synthesis is activated later during embryo-larval development. (See Domain of applicability)
Dose-Response Evidence: Empirical data is available from enough studies in animals treated with TPO inhibitors during development to make it readily accepted dogma that a dose-response relationship exists between TPO inhibition and serum TH concentrations. Again, these studies do not empirically measure TPO inhibition or decreased TH synthesis, but rely on the strong support of decades of mechanistic research in a variety of species of the causative relationship between these KEs. Examples of dose-responsive changes in TH concentrations following developmental exposure to TPO inhibitors include studies a variety of species, including: rodents (Blake and Henning, 1985; Goldey et al., 1995; Sawin et al., 1998); frogs (Tietge et al., 2013); fish tissue levels (Elsalini and Rohr, 2003.); and, chickens (Wishe et al., 1979). Computational modeling of the thyroid also provides evidence for the indirect dose-response relationship between these two KEs (e.g., Leonard et al., 2016; Fisher et a., 2013).
Uncertainties and InconsistenciesThere are no inconsistencies in this KER, but there are some uncertainties. The predominant uncertainty regarding the indirect key event relationship between inhibition of TPO activity and decreased serum T4 is the quantitative nature of this relationship, i.e., to what degree must TPO be inhibited in order to decrease serum T4 by a certain magnitude. Many animal (rat) studies typically employ relatively high exposures of TPO-inhibiting chemicals that result in hypothyroidism (severe decrements in T4 and T3). Thus, a dose-response relationship between TPO inhibition and decreased serum T4 is not typically defined. However, there are numerous publications demonstrating clear dose- and duration- dependent relationships between TPO inhibitors dose and reduced serum T3 and T4 in rodent models (see for example: Cooper et al., 1983; Hood et al., 1999; Goldey et al., 2005; Gilbert, 2011). The relationship between maternal and fetal levels of hormone following chemically-induced TPO inhibiton has not been well characterized and may differ based on kinetics. Reductions in serum TH in the fetus, in rat and human is derived a chemical’s effect on the maternal thyroid gland as well as the fetal thyroid gland.
Quantitative Understanding of the Linkage
Response-response relationshipThe indirect linkage between exposure to known TPO inhibitors and decreased serum TH has not been defined quantitatively. The two key event relationships that mediate this relationship (TPO inhibition leading to decreased TH synthesis, and decreased TH synthesis leading to decreased serum TH) have been incorporated into some quantitative models. A quantitative biologically-based dose-response model for iodine deficiency in the rat includes relationships between thyroidal T4 synthesis and serum T4 concentrations in developing rats Fisher et al. (2013). Ekerot et al. (2012) modeled TPO, T3, T4 and TSH in dogs and humans based on exposure to myeloperoxidase inhibitors that also inhibit TPO and was has recently adapted for rat (Leonard et al., 2016). While the original model predicted serum TH and TSH levels as a function of oral dose, it was not used to explicitly predict the relationship between serum hormones and TPO inhibition, or thyroidal hormone synthesis. Leonard et al. (2016) recently incorporated TPO inhibition into the model. Degon et al (2008) developed a human thyroid model that includes TPO but does not make quantitative prediction of organification changes due to inhibition of the TPO enzyme.
References
Ahmed OM, Abd El-Tawab SM, Ahmed RG. Effects of experimentally induced maternal hypothyroidism and hyperthyroidism on the development of rat offspring: I. The development of the thyroid hormones-neurotransmitters and adenosinergic system interactions. Int J Dev Neurosci. 2010 28(6):437-54
Axelstad M, Hansen PR, Boberg J, Bonnichsen M, Nellemann C, Lund SP, Hougaard KS, Hass U. Developmental neurotoxicity of propylthiouracil (PTU) in rats: relationship between transient hypothyroxinemia during development and long-lasting behavioural and functional changes. Toxicol Appl Pharmacol. 2008 232(1):1-13.
Blake HH, Henning SJ. Effect of propylthiouracil dose on serum thyroxine, growth, and weaning in young rats. Am J Physiol. 1985 248(5 Pt 2):R524-30.
Boas M, Feldt-Rasmussen U, Skakkebaek NE, Main KM. Environmental chemicals and thyroid function. Eur J Endocrinol. 2006 154:599-611.
Brucker-Davis F. Effects of environmental synthetic chemicals on thyroid function. Thyroid. 1998 8:827-56.
Calvo R, Obregón MJ, Ruiz de Oña C, Escobar del Rey F, Morreale de Escobar G. Congenital hypothyroidism, as studied in rats. Crucial role of maternal thyroxine but not of 3,5,3'-triiodothyronine in the protection of the fetal brain. J Clin Invest. 1990 Sep;86(3):889-99.
Chang J, Wang M, Gui W, Zhao Y, Yu L, Zhu G. 2012. Changes in thyroid hormone levels during zebrafish development. Zoological Science. 29(3):181-184.
Coady K, Marino T, Thomas J, Currie R, Hancock G, Crofoot J, McNalley L, McFadden L, Geter D, Klecka G. 2010. Evaluation of the amphibian metamorphosis assay: exposure to the goitrogen methimazole and the endogenous thyroid hormone L-thyroxine. Environmental toxicology and chemistry / SETAC. Apr;29:869-880.
Cooke PS, Kirby JD, Porcelli J. Increased testis growth and sperm production in adult rats following transient neonatal goitrogen treatment: optimization of the propylthiouracil dose and effects of methimazole. J Reprod Fertil. 1993 97(2):493-9
Cooper DS, Kieffer JD, Halpern R, Saxe V, Mover H, Maloof F, Ridgway EC (1983) Propylthiouracil (PTU) pharmacology in the rat. II. Effects of PTU on thyroid function. Endocrinology 113:921-928.
Crane, H.M., Pickford, D.B., Hutchinson, T.H., Brown, J.A., 2006. The effects of methimazole on development of the fathead minnow, pimephales promelas, from embryo to adult. Toxicological Sciences 93, 278-285.
Crofton KM Thyroid disrupting chemicals: mechanisms and mixtures. Int J Androl. 2008 31:209-23
Degon, M., Chipkin, S.R., Hollot, C.V., Zoeller, R.T., and Chait, Y. (2008). A computational model of the human thyroid. Mathematical Biosciences 212: 22–53.
Ekerot P, Ferguson D, Glämsta EL, Nilsson LB, Andersson H, Rosqvist S, Visser SA. Systems pharmacology modeling of drug-induced modulation of thyroid hormones in dogs and translation to human. Pharm Res. 2013 Jun;30(6):1513-24.
Elsalini OA, Rohr KB.: Phenylthiourea disrupts thyroid function in developing zebrafish. Dev Genes Evol 212, 593-8, 2003.
Emiliano, A.B., Governale, L., Parks, M., Cooper, D.S., 2010. Shifts in propylthiouracil and methimazole prescribing practices: antithyroid drug use in the United States from 1991 to 2008. J. Clin. Endocrinol. Metab. 95, 2227–2233.
Fisher JW, Li S, Crofton K, Zoeller RT, McLanahan ED, Lumen A, Gilbert ME. Evaluation of iodide deficiency in the lactating rat and pup using a biologically based dose-response model. Toxicol Sci. 2013 132(1):75-86.
Gilbert ME. 2011. Impact of low-level thyroid hormone disruption induced by propylthiouracil on brain development and function. Toxicol Sci. 124:432-445.
Goldey ES, Crofton KM. Thyroxine replacement attenuates hypothyroxinemia, hearing loss, and motor deficits following developmental exposure to Aroclor 1254 in rats. Toxicol Sci. 1998 Sep;45(1):94-105.
Goldey ES, Kehn LS, Rehnberg GL, Crofton KM. Effects of developmental hypothyroidism on auditory and motor function in the rat. Toxicol Appl Pharmacol. 1995 135(1):67-76.
Grommen, S.V., Iwasawa, A., Beck, V., Darras, V.M., De Groef, B., 2011. Ontogenic expression profiles of thyroid-specific genes in embryonic and hatching chicks. Domest. Anim. Endocrinol. 40, 10–18.
Hernandez-Mariano JA, Torres-Sanchez L, Bassol-Mayagoitia S, Escamilla-Nunez M, Cebrian ME, Villeda-Gutierrez EA, Lopez-Rodriguez G, Felix-Arellano EE, Blanco-Munoz J. 2017. Effect of exposure to p,p '-dde during the first half of pregnancy in the maternal thyroid profile of female residents in a mexican floriculture area. Environmental Research. 156:597-604.
Herwig A, Campbell G, Mayer CD, Boelen A, Anderson RA, Ross AW, Mercer JG, Barrett P. 2014. A thyroid hormone challenge in hypothyroid rats identifies T3 regulated genes in the hypothalamus and in models with altered energy balance and glucose homeostasis. Thyroid: Nov;24:1575-1593.
Hood A, Liu YP, Gattone VH, 2nd, Klaassen CD (1999) Sensitivity of thyroid gland growth to thyroid stimulating hormone (TSH) in rats treated with antithyroid drugs. Toxicol Sci 49:263-271.
Hurley PM. Mode of carcinogenic action of pesticides inducing thyroid follicular cell tumors in rodents. Environ Health Perspect. 1998 106:437-45.
Köhrle J. Environment and endocrinology: the case of thyroidology. Ann Endocrinol (Paris). 2008 69:116-22.
Lasley SM, Gilbert ME. Developmental thyroid hormone insufficiency reduces expression of brain-derived neurotrophic factor (BDNF) in adults but not in neonates. Neurotoxicol Teratol. 2011 33(4):464-72
Laurberg P, Andersen SL. 2014. Therapy of endocrine disease: antithyroid drug use in early pregnancy and birth defects: time windows of relative safety and high risk? Eur J Endocrinol. 2014 Jul;171(1):R13-20.
Leonard JA, Tan YM, Gilbert M, Isaacs K, El-Masri H. Estimating margin of exposure to thyroid peroxidase inhibitors using high-throughput in vitro data, high-throughput exposure modeling, and physiologically based pharmacokinetic/pharmacodynamic modeling. Toxicol Sci. 2016 151(1):57-70.
Liu XS, Cai Y, Wang Y, Xu SH, Ji K, Choi K. 2019. Effects of tris(1,3-dichloro-2-propyl) phosphate (tdcpp) and triphenyl phosphate (tpp) on sex-dependent alterations of thyroid hormones in adult zebrafish. Ecotoxicology and Environmental Safety. 170:25-32.
Männistö PT, Ranta T, Leppäluoto J. Effects of methylmercaptoimidazole (MMI), propylthiouracil (PTU), potassium perchlorate (KClO4) and potassium iodide (KI) on the serum concentrations of thyrotrophin (TSH) and thyroid hormones in the rat. Acta Endocrinol (Copenh). 1979 91(2):271-81.
Morreale de Escobar G, Obregon MJ, Escobar del Rey F (2004) Role of thyroid hormone during early brain development. Eur J Endocrinol 151 Suppl 3:U25-37.
Murk AJ, Rijntjes E, Blaauboer BJ, Clewell R, Crofton KM, Dingemans MM, Furlow JD, Kavlock R, Köhrle J, Opitz R, Traas T, Visser TJ, Xia M, Gutleb AC. Mechanism-based testing strategy using in vitro approaches for identification of thyroid hormone disrupting chemicals. Toxicol In Vitro. 2013 27:1320-46.
Nelson, K., Schroeder, A., Ankley, G., Blackwell, B., Blanksma, C., Degitz, S., Flynn, K., Jensen, K., Johnson, R., Kahl, M., Knapen, D., Kosian, P., Milsk, R., Randolph, E., Saari, T., Stinckens, E., Vergauwen, L., Villeneuve, D., 2016. Impaired anterior swim bladder inflation following exposure to the thyroid peroxidase inhibitor 2-mercaptobenzothiazole part I: Fathead minnow. Aquatic Toxicology 173, 192-203.
Opitz, R., Maquet, E., Zoenen, M., Dadhich, R., Costagliola, S., 2011. TSH Receptor Function Is Required for Normal Thyroid Differentiation in Zebrafish. Molecular Endocrinology 25, 1579-1599.
Pathak A, Sinha RA, Mohan V, Mitra K, Godbole MM. 2011. Maternal thyroid hormone before the onset of fetal thyroid function regulates reelin and downstream signaling cascade affecting neocortical neuronal migration. Cerebral Cortex. 21:11-21.
Pearce EN, Braverman LE. Environmental pollutants and the thyroid. Best Pract Res Clin Endocrinol Metab. 2009 23:801-1.
Power DM, Llewellyn L, Faustino M, Nowell MA, Bjornsson BT, Einarsdottir IE, Canario AV, Sweeney GE. 2001. Thyroid hormones in growth and development of fish. Comp Biochem Physiol C Toxicol Pharmacol. 130(4):447-459.
Rosebrough, R.W., Russell, B.A., McMurtry, J.P., 2006. Studies on doses of methimazole (MMI) and its administration regimen on broiler metabolism. Comp. Biochem. Physiol. A: Mol. Integr. Physiol. 143, 35–41.
Sack J, Weller A, Rigler O, Rozin A.A simple model for studying the correction of in utero hypothyroidism in the rat. Pediatr Res. 1995 37(4 Pt 1):497-501.
Sawin S, Brodish P, Carter CS, Stanton ME, Lau C. Development of cholinergic neurons in rat brain regions: dose-dependent effects of propylthiouracil-induced hypothyroidism. Neurotoxicol Teratol. 1998 20(6):627-35
Schwartz HL, Ross ME, Oppenheimer JH (1997) Lack of effect of thyroid hormone on late fetal rat brain development. Endocrinology 138:3119-3124.
Seed J, Carney EW, Corley RA, Crofton KM, DeSesso JM, Foster PM, Kavlock R, Kimmel G, Klaunig J, Meek ME, Preston RJ, Slikker W Jr, Tabacova S, Williams GM, Wiltse J, Zoeller RT, Fenner-Crisp P, Patton DE. Overview: Using mode of action and life stage information to evaluate the human relevance of animal toxicity data. Crit Rev Toxicol. 2005 35(8-9):664-72.
Shibutani M, Woo GH, Fujimoto H, Saegusa Y, Takahashi M, Inoue K, Hirose M, Nishikawa A. Assessment of developmental effects of hypothyroidism in rats from in utero and lactation exposure to anti-thyroid agents. Reprod Toxicol. 2009 Nov;28(3):297-307
Stinckens, E., Vergauwen, L., Blackwell, B.R., Anldey, G.T., Villeneuve, D.L., Knapen, D., 2020. Effect of Thyroperoxidase and Deiodinase Inhibition on Anterior Swim Bladder Inflation in the Zebrafish. Environmental Science & Technology 54, 6213-6223.
Stinckens, E., Vergauwen, L., Schroeder, A., Maho, W., Blackwell, B., Witters, H., Blust, R., Ankley, G., Covaci, A., Villeneuve, D., Knapen, D., 2016. Impaired anterior swim bladder inflation following exposure to the thyroid peroxidase inhibitor 2-mercaptobenzothiazole part II: Zebrafish. Aquatic Toxicology 173, 204-217.
Sundaresh V, Brito JP, Wang Z, Prokop LJ, Stan MN, Murad MH, Bahn RS. 2013. Comparative effectiveness of therapies for Graves' hyperthyroidism: a systematic review and network meta-analysis. The Journal of clinical endocrinology and metabolism. 98:3671-3677.
Taurog A. 2005. Hormone synthesis. In: Werner and Ingbar’s The Thyroid: A Fundamental and Clinical Text (Braverman LE, Utiger RD, eds). Philadelphia:Lippincott, Williams and Wilkins, 47–81
Taurog, a, Dorris, M. L., & Doerge, D. R. (1996). Mechanism of simultaneous iodination and coupling catalyzed by thyroid peroxidase. Archives of Biochemistry and Biophysics, Taurog A. Molecular evolution of thyroid peroxidase. Biochimie. 1999 May;81(5):557-62
Tietge JE, Butterworth BC, Haselman JT, Holcombe GW, Hornung MW, Korte JJ, Kosian PA, Wolfe M, Degitz SJ. Early temporal effects of three thyroid hormone synthesis inhibitors in Xenopus laevis. Aquat Toxicol. 2010 Jun 1;98(1):44-50
Tietge JE, Degitz SJ, Haselman JT, Butterworth BC, Korte JJ, Kosian PA, Lindberg-Livingston AJ, Burgess EM, Blackshear PE, Hornung MW. Inhibition of the thyroid hormone pathway in Xenopus laevis by 2-mercaptobenzothiazole. Aquat Toxicol. 2013 15;126:128-36
Tietge, J.E., Degitz, S.J., Haselman, J.T., Butterworth, B.C., Korte, J.J., Kosian, P.A., Lindberg-Livingston, A.J., Burgess, E.M., Blackshear, P.E., Hornung, M.W., 2012. Inhibition of the thyroid hormone pathway in Xenopus laevis by 2- mercaptobenzothiazole. Aquat. Toxicol. 126C, 128–136.
Trepanier, L.A., 2006. Medical management of hyperthyroidism. Clin. Tech. Small Anim. Pract. 21, 22–28.
U.S. Environmental Protection Agency. 2005. Guidance for Thyroid Assays in Pregnant Animals, Fetuses, and Postnatal Animals, and Adult Animals. Office of Pesticide Programs, Health Effects Division, Washington, DC
U.S. Environmental Protection Agency. 2009. Endocrine Disruptor Screening Program Test Guidelines OPPTS 890.1100: Amphibian Metamorphosis (Frog). Washington, DC.
Wabukebunoti MAN, Firling CE. 1983. The prehatching development of the thyroid-gland of the fathead minnow, pimephales-promelas (rafinesque). General and Comparative Endocrinology. 49(2):320-331.
Walter, K.M., Miller, G.W., Chen, X.P., Yaghoobi, B., Puschner, B., Lein, P.J., 2019. Effects of thyroid hormone disruption on the ontogenetic expression of thyroid hormone signaling genes in developing zebrafish (Danio rerio). General and Comparative Endocrinology 272, 20-32.
Webster GM, Venners SA, Mattman A, Martin JW. 2014. Associations between perfluoroalkyl acids (pfass) and maternal thyroid hormones in early pregnancy: A population-based cohort study. Environmental Research. 133:338-347.
Wishe H I, Rolle-Getz G K, and Goldsmith E D.: The effects of aminotriazole (ATZ) on the thyroid gland and the development of the white leghorn chick. Growth 43, 238-251, 1979
Wu S, Tan G, Dong X, Zhu Z, Li W, Lou Z, Chai Y. 2013. Metabolic profiling provides a system understanding of hypothyroidism in rats and its application. PloS one.8:e55599.
Zoeller RT, Crofton KM (2005) Mode of action: developmental thyroid hormone insufficiency--neurological abnormalities resulting from exposure to propylthiouracil. Crit Rev Toxicol 35:771-781.