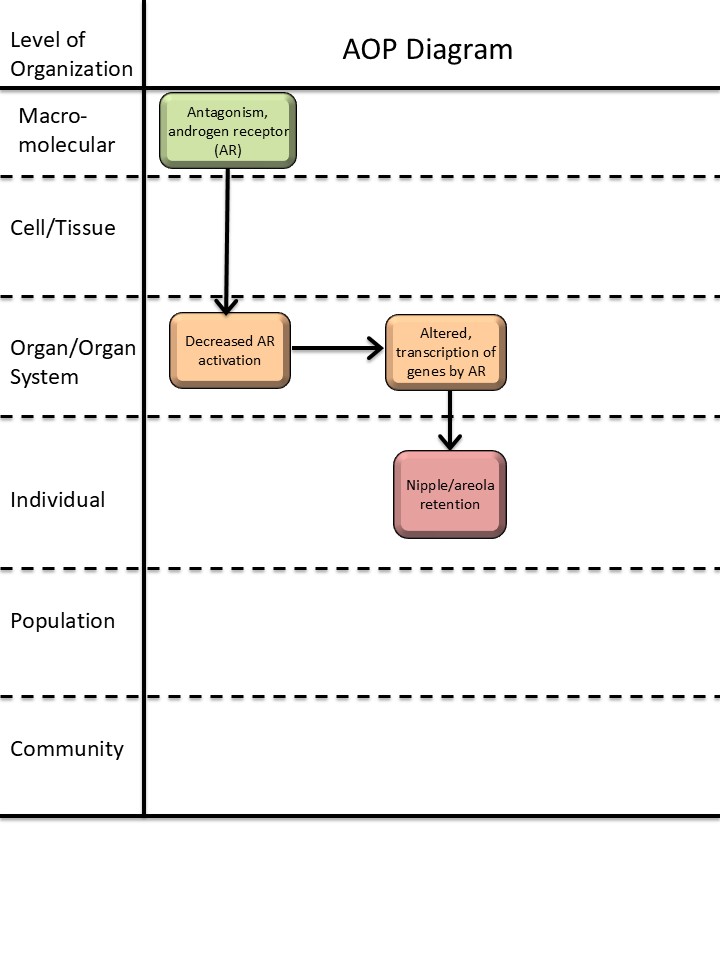
AOP ID and Title:
Graphical Representation
Status
| Author status | OECD status | OECD project | SAAOP status |
|---|---|---|---|
| Under development: Not open for comment. Do not cite | Under Development | 1.108 | Included in OECD Work Plan |
Abstract
This AOP links androgen receptor (AR) antagonism during fetal life with nipple/areola retention (NR) in male rodent offspring. NR, measured around 2 weeks postpartum I laboratory mice and rats, is a marker for feminization of male offspring.
The AR is a nuclear receptor involved in the transcriptional regulation of various target genes during development and adulthood across species. Its main ligands are testosterone and dihydrotestosterone (DHT). Under normal physiological conditions, testosterone, produced mainly by the testes, is converted by 5α-reductase to DHT locally in tissues; in turn DHT binds AR and activates downstream target genes. AR signaling is necessary for normal masculinization of the developing fetus, and AR action in male rodents signals the nipple anlagen to regress, leaving males with no nipples.
The key events in this pathway are fetal antagonism of the AR in target cells of the nipple anlagen, which leads to inactivation of the AR and failure to suppress development of the nipples, causing retention of nipples, visible postnatally in male offspring. In this instance, the local levels of testosterone or DHT may be normal but prevented from binding to the AR. Downstream of a reduction in AR activation, the molecular mechanisms of nipple retention are unclear, highlighting a knowledge gap in this AOP and potential for further development.
The confidence in each of the KERs comprising the AOP is judged as high, with both high biological plausibility and high confidence in empirical evidence. The mechanistic link between KE-286 (‘altered, transcription of genes by AR’) and AO-1786 (‘Increase, Nipple retention’) is not established, but given the high confidence in the KERs, the overall confidence in the AOP is judged as high.
The AOP supports the regulatory application of NR as a measure of endocrine disruption relevant for human health and the use of NR as an indicator of anti-androgenicity in environmentally relevant species. Even though NR cannot be directly translated to a human endpoint, the AOP is considered human relevant since NR is a clear readout of reduced androgen action and masculinization during development and is considered an ‘adverse outcome’ in OECD test guidelines (TG 443, TG 421, TG 422). The AOP also holds utility for informing on anti-androgenicity more generally, as this modality is highly relevant across mammalian species and vertebrates more broadly due to the conserved nature of the AR and its implication in sexual differentiation across species.
Background
This AOP is a part of an AOP network for reduced androgen receptor activation leading to retention of nipples/areolas in male offspring. The other AOPs in this network are AOP-575 (‘Decreased intratesticular testosterone leading to increased nipple retention (NR) in male (mouse and rat) offspring’) and AOP-576 (‘5α-reductase inhibition leading to increased nipple retention (NR) in male (mouse and rat) offspring’). The purpose of the AOP network is to organize the well-established evidence for anti-androgenic mechanisms-of-action leading to increased NR. It can be used in identification and assessment of endocrine disruptors and to inform predictive toxicology, identification of knowledge gaps for investigation and method development.
This work received funding from the European Food and Safety Authority (EFSA) under Grant agreement no. GP/EFSA/PREV/2022/01.
Summary of the AOP
Events
Molecular Initiating Events (MIE), Key Events (KE), Adverse Outcomes (AO)
| Sequence | Type | Event ID | Title | Short name |
|---|---|---|---|---|
| MIE | 26 | Antagonism, Androgen receptor | Antagonism, Androgen receptor | |
| KE | 1614 | Decrease, androgen receptor activation | Decrease, AR activation | |
| KE | 286 | Altered, Transcription of genes by the androgen receptor | Altered, Transcription of genes by the AR | |
| AO | 1786 | Nipple retention (NR), increased | nipple retention, increased |
Key Event Relationships
| Upstream Event | Relationship Type | Downstream Event | Evidence | Quantitative Understanding |
|---|---|---|---|---|
| Antagonism, Androgen receptor | adjacent | Decrease, androgen receptor activation | High | |
| Decrease, androgen receptor activation | adjacent | Altered, Transcription of genes by the androgen receptor | High | |
| Antagonism, Androgen receptor | non-adjacent | Nipple retention (NR), increased | High | |
| Decrease, androgen receptor activation | non-adjacent | Nipple retention (NR), increased | High |
Stressors
| Name | Evidence |
|---|---|
| Flutamide | |
| Vinclozolin | |
| Procymidone |
Overall Assessment of the AOP
Domain of Applicability
Life Stage Applicability| Life Stage | Evidence |
|---|---|
| Foetal | High |
| Sex | Evidence |
|---|---|
| Male | High |
The upstream part of the AOP has a broad applicability domain, but the downstream KERs-2133 (Antagonism, AR, leads to increased nipple retention) and KER-3348 (Decrease, AR activation, leads to increased nipple retention) are considered only directly applicable to male rodents (evidence primarily from laboratory rats and mice) during fetal life, restricting the taxonomic applicability of the AOP. Although NR is a feature having been investigated in laboratory rats and mice, it is biologically plausible that the AOP is applicable to other rodent species. The process of retention of nipples by disruption of androgen programming happens in the fetal life stage, but the AO is detected postnatally. In the males of mice and rats, the nipple anlagen are programmed during fetal development by androgens to regress, leading to no visible nipples in males postnatally, while females exhibit nipples. This AOP only contains empirical evidence for the applicability to male rats, but the AOP is considered equally applicable to male mice, as these also normally exhibit nipple regression stimulated by androgens. Moreover, the AOP is indirectly relevant for other taxa, including humans, as nipple retention in male rodents indicates a reduction in fetal masculinization. Nipple retention is therefore included as a mandatory endpoint in multiple OECD Test Guideline studies for developmental and reproductive toxicity and is considered applicable as an adverse outcome to set NOAELs and LOAELs of substances in human health risk assessments.
Essentiality of the Key Events
|
Evidence |
Uncertainties, inconsistencies and contradictory evidence |
|
|
MIE-26 Antagonism, AR receptor
HIGH: This MIE is usually measured in vitro, whereas the downstream events in the AOP are, in most cases measured in vivo. Canonical knowledge of normal male reproductive development provides strong support for essentiality, along with AR knockout models.
|
Biological plausibility provides strong support for the essentiality of this event, as androgens, acting through AR, are the primary drivers of regression of nipple anlagen in male rat and mice embryos (Imperato-McGinley et al., 1986; Kratochwil, 1977; Kratochwil & Schwartz, 1976).
Indirect evidence of the impact of AR antagonism (MIE-26) in vitro on AR activity in vitro: • Several chemical substances, including flutamide and vinclozolin, are known AR antagonists and have been shown to decrease AR activity in vitro (Pedersen et al., 2022; Sonneveld et al., 2004).
Indirect evidence of the impact of AR antagonism (MIE-26) in vivo on increased nipple retention (AO-1786): • Rat in vivo exposure to vinclozolin, procymidone and flutamide, which are known AR antagonists, leads to increased nipple retention in offspring (see KER-3348).
Direct evidence of the impact of AR antagonism (MIE-26) in vivo on increased nipple retention (AO-1786): • Male Tfm mutant mice, which are insensitive to androgens and believed to be so due to a nonfunctional androgen receptor, present with retained nipples (Kratochwil & Schwartz, 1976) |
|
|
KE-1614 Decreased, AR activation
HIGH: There is experimental evidence from mutant mice insensitive to androgens showing that the AR is essential for nipple retention in male offspring. There is also evidence from exposure studies in animals that substances antagonizing AR induce nipple retention in male pups. |
Biological plausibility provides strong support for the essentiality of this event, as AR activation is critical for normal regression of nipple anlagen in male embryos.
Indirect evidence of the impact of decreased AR activation (KE-1614) on altered gene transcription by AR (KE-286): • Exposure to known anti-androgenic chemicals induces a changed gene expression pattern, e.g. in neonatal pig ovaries (Knapczyk-Stwora et al., 2019).
Direct evidence of the impact of decreased AR activation (KE-1614) on altered gene transcription by AR (KE-286): • Male AR KO mice have altered gene expression patterns in a broad range of organs (refer to KER-2124).
Indirect evidence of the impact of decreased AR activation (KE-1614) on increased nipple retention (AO-1786): • Rat in vivo exposure to vinclozolin, procymidone and flutamide, which are known AR antagonists, leads to increased nipple retention in offspring (see KER-3348).
Direct evidence of the impact of decreased AR activation (KE-1614) on increased nipple retention (AO-1786): • Male Tfm mutant mice, which are insensitive to androgens and believed to be so due to a nonfunctional androgen receptor, present with retained nipples (Kratochwil & Schwartz, 1976) |
|
|
KE-286 Altered, trans. of genes by AR
LOW: Strongest support for essentiality comes from biological plausibility. However, exact transcriptional effects and causality remain to be fully characterized. |
Biological plausibility provides support for the essentiality of this event. AR is a nuclear receptor and transcription factor regulating transcription of genes, and androgens, acting through AR, are essential for normal regression of nipple anlagen in male fetuses. |
There are currently no AR-responsive genes proven to be causally involved in nipple retention, and it is known that AR can also signal through non-genomic actions (Leung & Sadar, 2017). |
|
Direct evidence |
Indirect evidence |
Contradictory evidence |
Overall essentiality assessment |
|
|
MIE-26 |
*** |
** |
|
High |
|
KE-1614 |
*** |
*** |
|
High |
|
KE-286 |
|
|
|
Low (biological plausibility) |
*Low level of evidence (some support for essentiality), ** Intermediate level of evidence (evidence for impact on one or more downstream KEs), ***High level of evidence (evidence for impact on AO).
Weight of Evidence Summary
The confidence in each of the KERs comprising the AOP is judged as high, with both high biological plausibility and high confidence in empirical evidence. The mechanistic link between KE-286 (‘altered, transcription of genes by AR’) and AO-1786 (‘Increase, Nipple retention’) is not established, but given the high confidence in the KERs, the overall confidence in the AOP is judged as high.
|
KER |
Biological Plausibility |
Empirical Evidence |
Rationale |
|
KER-2130 Antagonism, AR leads to decrease, AR activation |
High |
High (canonical) |
It is well established that antagonism of the AR leads to decreased AR activity. Direct evidence for this KER is not possible since KE-1614 can currently not be measured and is considered an in vivo effect. Indirect evidence using proxy read-outs of AR activation, either in vitro or in vivo, strongly supports the relationship (Draskau et al., 2024) |
|
KER-2124 Decrease, AR activation leads to altered, transcription of genes by AR |
High |
High (canonical) |
It is well established that the AR regulates gene transcription. In vivo animal studies and human genomic profiling show tissue-specific changes to gene expression upon disruption of AR. |
|
KER-2133 Antagonism, AR leads to increased nipple retention |
High |
High |
It is well established that androgens drive the regression of nipple anlagen in male rat and mouse fetuses through interaction with the AR receptor. The biological plausibility is high, and so is the empirical evidence, which includes numerous rat studies showing increased nipple retention in male offspring after exposure to well-known anti-androgens. |
|
KER-3348 Decrease, AR activation leads to increased nipple retention. |
High |
High |
It is well established that activation of AR drives the regression of nipple anlagen in males. The empirical evidence includes numerous in vivo toxicity studies showing that decreased AR activation leads to increased NR in male offspring, with few inconsistencies. The empirical evidence combined with theoretical considerations provides some support for dose, temporal, and incidence concordance for the KER, although this evidence is weak and indirect. |
Quantitative Consideration
The quantitative understanding of the AOP is limited. A key difficulty lies in the challenge of extrapolating from in vitro to in vivo events since these cannot be captured within the same experimental framework. Specifically, MIE-26 is evaluated in vitro, while both the AO (NR) and KE-1614 are in vivo endpoints. KE-1614 pertains to AR activation in vivo - currently lacking viable methods for direct measurement.
The difficulties with in vitro-to-in vivo potency extrapolation from studies were exemplified by a comparison of the effects of pyrifluquinazon and bisphenol C in vitro and in utero. In vitro, bisphenol C antagonized the androgen receptor with a much higher potency than pyrifluquinazon, but in vivo the potencies were reversed with pyrifluquinazon exposure leading to NR at lower exposure levels than bisphenol C (Gray et al., 2019).
Considerations for Potential Applications of the AOP (optional)
The AOP supports the regulatory application of NR as a measure of endocrine disruption relevant for human health and the use of NR as an indicator of anti-androgenicity in mammals and other vertebrates in the environment.
NR is a mandatory endpoint in multiple OECD test guidelines, including TG 443 (extended one-generation reproductive toxicity study) and TGs 421/422 (reproductive toxicity screening studies) (OECD 2025a; OECD 2025b; OECD 2025c). NR can contribute to establishing a No Observed Adverse Effect Level (NOAEL), as outlined in OECD guidance documents No. 43 and 151 (OECD 2008; OECD 2013). The ability to derive a NOAEL for increased NR in male rodent offspring, which can serve as a point of departure for determining human safety thresholds, underscores the regulatory significance of this AOP.
The AOP also holds utility for informing on anti-androgenicity more generally, as this modality is highly relevant across mammalian species (Schwartz et al., 2021) and vertebrates more broadly due to the conserved nature of the AR and its implication in sexual differentiation across species (Ogino et al., 2023).
References
Chamberlain, N. L., Driver, E. D., & Miesfeld, R. L. (1994). The length and location of CAG trinucleotide repeats in the androgen receptor N-terminal domain affect transactivation function. Nucleic Acids Research, 22(15), 3181–3186. https://doi.org/10.1093/nar/22.15.3181
Draskau, M. K., Rosenmai, A. K., Bouftas, N., Johansson, H. K. L., Panagiotou, E. M., Holmer, M. L., Elmelund, E., Zilliacus, J., Beronius, A., Damdimopoulou, P., van Duursen, M., & Svingen, T. (2024). AOP Report: An Upstream Network for Reduced Androgen Signaling Leading to Altered Gene Expression of Androgen Receptor–Responsive Genes in Target Tissues. Environmental Toxicology and Chemistry, 43(11), 2329–2337. https://doi.org/10.1002/etc.5972
Gray, L. E., Furr, J. R., Conley, J. M., Lambright, C. S., Evans, N., Cardon, M. C., Wilson, V. S., Foster, P. M., & Hartig, P. C. (2019). A Conflicted Tale of Two Novel AR Antagonists in Vitro and in Vivo: Pyrifluquinazon Versus Bisphenol C. Toxicological Sciences, 168(2), 632–643. https://doi.org/10.1093/toxsci/kfz010
Holmer ML, Zilliacus J, Draskau MK, Hlisníková H, Beronius A, Svingen T. Methodology for developing data-rich Key Event Relationships for Adverse Outcome Pathways exemplified by linking decreased androgen receptor activity with decreased anogenital distance. Reprod Toxicol. 2024 Sep;128:108662. doi: 10.1016/j.reprotox.2024.108662 . Epub 2024 Jul 8. PMID: 38986849.
Imperato-McGinley J, Binienda Z, Gedney J, & Vaughan ED Jr. (1986). Nipple differentiation in fetal male rats treated with an inhibitor of the enzyme 5 alpha-reductase: definition of a selective role for dihydrotestosterone. Endocrinology, 118(1), 132–137. https://doi.org/10.1210/endo-118-1-132
Knapczyk-Stwora, K., Nynca, A., Ciereszko, R. E., Paukszto, L., Jastrzebski, J. P., Czaja, E., Witek, P., Koziorowski, M., & Slomczynska, M. (2019). Flutamide-induced alterations in transcriptional profiling of neonatal porcine ovaries. Journal of Animal Science and Biotechnology, 10(1), 35. https://doi.org/10.1186/s40104-019-0340-y
Kratochwil, K. (1977). Development and loss of androgen responsiveness in the embryonic rudiment of the mouse mammary gland. Developmental Biology, 61(2), 358–365. https://doi.org/10.1016/0012-1606(77)90305-0
Kratochwil, K., & Schwartz, P. (1976). Tissue interaction in androgen response of embryonic mammary rudiment of mouse: identification of target tissue for testosterone. Proceedings of the National Academy of Sciences, 73(11), 4041–4044. https://doi.org/10.1073/pnas.73.11.4041
Leung, J. K., & Sadar, M. D. (2017). Non-Genomic Actions of the Androgen Receptor in Prostate Cancer. Frontiers in Endocrinology, 8. https://doi.org/10.3389/fendo.2017.00002
OECD (2013), Guidance Document Supporting OECD Test Guideline 443 on the Extended One-Generational Reproductive Toxicity Test, OECD Series on Testing and Assessment, No. 151, OECD Publishing, Paris, ENV/JM/MONO(2013)10
OECD (2025a). Test No. 421: Reproduction/Developmental Toxicity Screening Test. https://doi.org/10.1787/9789264264380-en
OECD (2025b). Test No. 422: Combined Repeated Dose Toxicity Study with the Reproduction/Developmental Toxicity Screening Test. https://doi.org/10.1787/9789264264403-en
OECD (2025c). Test No. 443: Extended One-Generation Reproductive Toxicity Study. https://doi.org/10.1787/9789264185371-en
Ogino, Y., Ansai, S., Watanabe, E. et al. Evolutionary differentiation of androgen receptor is responsible for sexual characteristic development in a teleost fish. Nat Commun 14, 1428 (2023). https://doi.org/10.1038/s41467-023-37026-6
Pedersen, E. B., Christiansen, S., & Svingen, T. (2022). AOP key event relationship report: Linking androgen receptor antagonism with nipple retention. In Current Research in Toxicology (Vol. 3). Elsevier B.V. https://doi.org/10.1016/j.crtox.2022.100085
Schwartz, C. L., Christiansen, S., Hass, U., Ramhøj, L., Axelstad, M., Löbl, N. M., & Svingen, T. (2021). On the Use and Interpretation of Areola/Nipple Retention as a Biomarker for Anti-androgenic Effects in Rat Toxicity Studies. In Frontiers in Toxicology (Vol. 3). Frontiers Media S.A. https://doi.org/10.3389/ftox.2021.730752
Sonneveld, E., Jansen, H. J., Riteco, J. A., Brouwer, A., & van der Burg, B. (2004). Development of Androgen- and Estrogen-Responsive Bioassays, Members of a Panel of Human Cell Line-Based Highly Selective Steroid-Responsive Bioassays. Toxicological Sciences, 83(1), 136–148. https://doi.org/10.1093/toxsci/kfi005
Svingen, T., Villeneuve, D. L., Knapen, D., Panagiotou, E. M., Draskau, M. K., Damdimopoulou, P., & O’Brien, J. M. (2021). A Pragmatic Approach to Adverse Outcome Pathway Development and Evaluation. Toxicological Sciences, 184(2), 183–190. https://doi.org/10.1093/toxsci/kfab113
Tut, T. G., Ghadessy, F. J., Trifiro, M. A., Pinsky, L., & Yong, E. L. (1997). Long Polyglutamine Tracts in the Androgen Receptor Are Associated with Reduced Trans -Activation, Impaired Sperm Production, and Male Infertility 1. The Journal of Clinical Endocrinology & Metabolism, 82(11), 3777–3782. https://doi.org/10.1210/jcem.82.11.4385
Wolf, C., Lambright, C., Mann, P., Price, M., Cooper, R. L., Ostby, J., & Earl Gray, L. J. (1999). Administration of potentially antiandrogenic pesticides (procymidone, linuron, iprodione, chlozolinate, p,p-DDE, and ketoconazole) and toxic substances (dibutyl-and diethylhexyl phthalate, PCB 169, and ethane dimethane sulphonate) during sexual differentiation produces diverse profiles of reproductive malformations in the male rat. Toxicology and Industrial Health, 15, 94–118. www.stockton-press.co.uk
You L, Casanova M, Archibeque-Engle S, Sar M, Fan LQ, & Heck HA. (1998). Impaired male sexual development in perinatal Sprague-Dawley and Long-Evans hooded rats exposed in utero and lactationally to p,p’-DDE. Toxicological Sciences : An Official Journal of the Society of Toxicology, 45(2), 162–173. https://doi.org/10.1093/toxsci/45.2.162
Appendix 1
List of MIEs in this AOP
Event: 26: Antagonism, Androgen receptor
Short Name: Antagonism, Androgen receptor
Key Event Component
| Process | Object | Action |
|---|---|---|
| androgen receptor activity | androgen receptor | decreased |
AOPs Including This Key Event
| AOP ID and Name | Event Type |
|---|---|
| Aop:306 - Androgen receptor (AR) antagonism leading to short anogenital distance (AGD) in male (mammalian) offspring | MolecularInitiatingEvent |
| Aop:344 - Androgen receptor (AR) antagonism leading to nipple retention (NR) in male (mammalian) offspring | MolecularInitiatingEvent |
| Aop:345 - Androgen receptor (AR) antagonism leading to decreased fertility in females | MolecularInitiatingEvent |
| Aop:372 - Androgen receptor antagonism leading to testicular cancer | MolecularInitiatingEvent |
| Aop:477 - Androgen receptor (AR) antagonism leading to hypospadias in male (mammalian) offspring | MolecularInitiatingEvent |
| Aop:476 - Adverse Outcome Pathways diagram related to PBDEs associated male reproductive toxicity | MolecularInitiatingEvent |
| Aop:19 - Androgen receptor antagonism leading to adverse effects in the male foetus (mammals) | MolecularInitiatingEvent |
| Aop:595 - Nanoplastic effect | MolecularInitiatingEvent |
Stressors
| Name |
|---|
| Mercaptobenzole |
| Triticonazole |
| Flusilazole |
| Epoxiconazole |
| Prochloraz |
| Propiconazole |
| Tebuconazole |
| Flutamide |
| Cyproterone acetate |
| Vinclozolin |
Biological Context
| Level of Biological Organization |
|---|
| Molecular |
Cell term
| Cell term |
|---|
| eukaryotic cell |
Domain of Applicability
Taxonomic Applicability| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| mammals | mammals | High | NCBI |
| Life Stage | Evidence |
|---|---|
| During development and at adulthood | High |
| Sex | Evidence |
|---|---|
| Mixed | High |
Both the DNA-binding and ligand-binding domains of the AR are highly evolutionary conserved, whereas the transactivation domain show more divergence which may affect AR-mediated gene regulation across species (Davey & Grossmann, 2016). Despite certain inter-species differences, AR function mediated through gene expression is highly conserved, with mutations studies from both humans and rodents showing strong correlation for AR-dependent development and function (Walters et al, 2010).
This KE is applicable for both sexes, across developmental stages into adulthood, in numerous cells and tissues and across mammalian taxa. It is, however, acknowledged that this KE most likely has a much broader domain of applicability extending to non-mammalian vertebrates. AOP developers are encouraged to add additional relevant knowledge to expand on the applicability to also include other vertebrates.
Key Event Description
The androgen receptor (AR) and its function
The AR is a ligand-activated transcription factor belonging to the steroid hormone nuclear receptor family (Davey & Grossmann, 2016). The AR has three domains: the N-terminal domain, the DNA-binding domain and the ligand-binding domain, with the latter being most evolutionary conserved. Testosterone (T) and the more biologically active dihydrotestosterone (DHT) are endogenous ligands for the AR (MacLean et al, 1993; MacLeod et al, 2010; Schwartz et al, 2019). In teleost fishes, 11-ketotestosterone is the second main ligand (Schuppe et al, 2020). Human AR mutations and mouse knock-out models have established a pivotal role for the AR in masculinization and spermatogenesis (Walters et al, 2010). Apart from the essential role for AR in male reproductive development and function (Walters et al, 2010), the AR is also expressed in many other tissues and organs such as bone, muscles, ovaries, and the immune system (Rana et al, 2014).
AR antagonism as Key Event
The main function of the AR is to activate gene transcription in cells. Canonical signaling occurs by ligands (androgens) binding to AR in the cytoplasm which results in translocation to the cell nucleus, receptor dimerization and binding to specific regulatory DNA sequences (Heemers & Tindall, 2007). The gene targets regulated by AR activation depends on cell/tissue type and what stage of development activation occur, and is, for instance, dependent on available co-factors. Apart from the canonical signaling pathway, AR can also initiate cytoplasmic signaling pathways with other functions than the nuclear pathway, for instance rapid change in cell function by ion transport changes (Heinlein & Chang, 2002) and association with Src kinase to activate MAPK/ERK signaling and activation of the PI3K/Akt pathway (Leung & Sadar, 2017).
How it is Measured or Detected
AR antagonism can be measured in vitro by transient or stable transactivation assays to evaluate nuclear receptor activation. There is already a validated test guideline for AR (ant)agonism adopted by the OECD, Test No. 458: Stably Transfected Human Androgen Receptor Transcriptional Activation Assay for Detection of Androgenic Agonist and Antagonist Activity of Chemicals (OECD, 2016). This test guideline contains three different methods. More information on limitations, advantages, protocols, and availability and description of cells are given in the test guideline.
Besides these validated methods, other transiently or stably transfected reporter cell lines are available as well as yeast based systems (Campana et al, 2015; Körner et al, 2004). AR nuclear translocation can be monitored by various assays (Campana et al 2015), for example by monitoring fluorescent rat AR movement in living cells (Tyagi et al 2020), with several human AR translocation assays being commercially available; e.g. Fluorescent AR Nuclear Translocation Assay (tGFP-hAR/HEK293) or Human Androgen NHR Cell Based Antagonist Translocation LeadHunter Assay.
Additional information on AR interaction can be obtained employing competitive AR binding assays (Freyberger et al 2010, Shaw et al 2018), which can also inform on relative potency of the compounds, though not on downstream effect of the AR binding.
The recently developed AR dimerization assay provides an assay with an improved ability to measure potential stressor-mediated disruption of dimerization/activation (Lee et al, 2021).
References
Campana C, Pezzi V, Rainey WE (2015) Cell based assays for screening androgen receptor ligands. Semin Reprod Med 33: 225-234.
Freyberger A, Weimer M, Tran HS, Ahr HJ. Assessment of a recombinant androgen receptor binding assay: initial steps towards validation. Reprod Toxicol. 2010 Aug;30(1):2-8. doi: 10.1016/j.reprotox.2009.10.001. Epub 2009 Oct 13. PMID: 19833195.
OECD (2022). Test No. 251: Rapid Androgen Disruption Activity Reporter (RADAR) assay. Paris: OECD Publishing doi:10.1787/da264d82-en.
Shaw J, Leveridge M, Norling C, Karén J, Molina DM, O'Neill D, Dowling JE, Davey P, Cowan S, Dabrowski M, Main M, Gianni D. Determining direct binders of the Androgen Receptor using a high-throughput Cellular Thermal Shift Assay. Sci Rep. 2018 Jan 9;8(1):163. doi: 10.1038/s41598-017-18650-x. PMID: 29317749; PMCID: PMC5760633.
Tyagi RK, Lavrovsky Y, Ahn SC, Song CS, Chatterjee B, Roy AK (2000) Dynamics of intracellular movement and nucleocytoplasmic recycling of the ligand-activated androgen receptor in living cells. Mol Endocrinol 14: 1162-1174
List of Key Events in the AOP
Event: 1614: Decrease, androgen receptor activation
Short Name: Decrease, AR activation
Key Event Component
| Process | Object | Action |
|---|---|---|
| androgen receptor activity | androgen receptor | decreased |
AOPs Including This Key Event
Biological Context
| Level of Biological Organization |
|---|
| Tissue |
Domain of Applicability
Taxonomic Applicability| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| mammals | mammals | High | NCBI |
| Life Stage | Evidence |
|---|---|
| During development and at adulthood | High |
| Sex | Evidence |
|---|---|
| Mixed | High |
This KE is considered broadly applicable across mammalian taxa as all mammals express the AR in numerous cells and tissues where it regulates gene transcription required for developmental processes and functions. It is, however, acknowledged that this KE most likely has a much broader domain of applicability extending to non-mammalian vertebrates. AOP developers are encouraged to add additional relevant knowledge to expand on the applicability to also include other vertebrates.
Key Event Description
This KE refers to decreased activation of the androgen receptor (AR) as occurring in complex biological systems such as tissues and organs in vivo. It is thus considered distinct from KEs describing either blocking of AR or decreased androgen synthesis.
The AR is a nuclear transcription factor with canonical AR activation regulated by the binding of the androgens such as testosterone or dihydrotestosterone (DHT). Thus, AR activity can be decreased by reduced levels of steroidal ligands (testosterone, DHT) or the presence of compounds interfering with ligand binding to the receptor (Davey & Grossmann, 2016; Gao et al., 2005).
In the inactive state, AR is sequestered in the cytoplasm of cells by molecular chaperones. In the classical (genomic) AR signaling pathway, AR activation causes dissociation of the chaperones, AR dimerization and translocation to the nucleus to modulate gene expression. AR binds to the androgen response element (ARE) (Davey & Grossmann, 2016; Gao et al., 2005). Notably, for transcriptional regulation the AR is closely associated with other co-factors that may differ between cells, tissues and life stages. In this way, the functional consequence of AR activation is cell- and tissue-specific. This dependency on co-factors such as the SRC proteins also means that stressors affecting recruitment of co-activators to AR can result in decreased AR activity (Heinlein & Chang, 2002).
Ligand-bound AR may also associate with cytoplasmic and membrane-bound proteins to initiate cytoplasmic signaling pathways with other functions than the nuclear pathway. Non-genomic AR signaling includes association with Src kinase to activate MAPK/ERK signaling and activation of the PI3K/Akt pathway. Decreased AR activity may therefore be a decrease in the genomic and/or non-genomic AR signaling pathways (Leung & Sadar, 2017).
How it is Measured or Detected
This KE specifically focuses on decreased in vivo activation, with most methods that can be used to measure AR activity carried out in vitro. They provide indirect information about the KE and are described in lower tier MIE/KEs (see for example MIE/KE-26 for AR antagonism, KE-1690 for decreased T levels and KE-1613 for decreased dihydrotestosterone levels). Assays may in the future be developed to measure AR activation in mammalian organisms.
References
Davey, R. A., & Grossmann, M. (2016). Androgen Receptor Structure, Function and Biology: From Bench to Bedside. The Clinical Biochemist. Reviews, 37(1), 3–15.
Gao, W., Bohl, C. E., & Dalton, J. T. (2005). Chemistry and structural biology of androgen receptor. Chemical Reviews, 105(9), 3352–3370. https://doi.org/10.1021/cr020456u
Heinlein, C. A., & Chang, C. (2002). Androgen Receptor (AR) Coregulators: An Overview. https://academic.oup.com/edrv/article/23/2/175/2424160
Leung, J. K., & Sadar, M. D. (2017). Non-Genomic Actions of the Androgen Receptor in Prostate Cancer. Frontiers in Endocrinology, 8. https://doi.org/10.3389/fendo.2017.00002
OECD (2022). Test No. 251: Rapid Androgen Disruption Activity Reporter (RADAR) assay. Paris: OECD Publishing doi:10.1787/da264d82-en.
|
|
|
Event: 286: Altered, Transcription of genes by the androgen receptor
Short Name: Altered, Transcription of genes by the AR
Key Event Component
| Process | Object | Action |
|---|---|---|
| regulation of gene expression | androgen receptor | decreased |
AOPs Including This Key Event
Stressors
| Name |
|---|
| Bicalutamide |
| Cyproterone acetate |
| Epoxiconazole |
| Flutamide |
| Flusilazole |
| Prochloraz |
| Propiconazole |
| Stressor:286 Tebuconazole |
| Triticonazole |
| Vinclozalin |
Biological Context
| Level of Biological Organization |
|---|
| Tissue |
Domain of Applicability
Taxonomic Applicability| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| mammals | mammals | High | NCBI |
| Life Stage | Evidence |
|---|---|
| During development and at adulthood | High |
| Sex | Evidence |
|---|---|
| Mixed | High |
Both the DNA-binding and ligand-binding domains of the AR are highly evolutionary conserved, whereas the transactivation domain show more divergence, which may affect AR-mediated gene regulation across species (Davey and Grossmann 2016). Despite certain inter-species differences, AR function mediated through gene expression is highly conserved, with mutation studies from both humans and rodents showing strong correlation for AR-dependent development and function (Walters et al. 2010).
This KE is considered broadly applicable across mammalian taxa, sex and developmental stages, as all mammals express the AR in numerous cells and tissues where it regulates gene transcription required for developmental processes and function. It is, however, acknowledged that this KE most likely has a much broader domain of applicability extending to non-mammalian vertebrates. AOP developers are encouraged to add additional relevant knowledge to expand on the applicability to also include other vertebrates.
Key Event Description
This KE refers to transcription of genes by the androgen receptor (AR) as occurring in complex biological systems such as tissues and organs in vivo. Rather than measuring individual genes, this KE aims to capture patterns of effects at transcriptome level in specific target cells/tissues. In other words, it can be replaced by specific KEs for individual adverse outcomes as information becomes available, for example the transcriptional toxicity response in prostate tissue for AO: prostate cancer, perineum tissue for AO: reduced AGD, etc. AR regulates many genes that differ between tissues and life stages and, importantly, different gene transcripts within individual cells can go in either direction since AR can act as both transcriptional activator and suppressor. Thus, the ‘directionality’ of the KE cannot be either reduced or increased, but instead describe an altered transcriptome.
The Androgen Receptor and its function
The AR belongs to the steroid hormone nuclear receptor family. It is a ligand-activated transcription factor with three domains: the N-terminal domain, the DNA-binding domain, and the ligand-binding domain with the latter being the most evolutionary conserved (Davey and Grossmann 2016). Androgens (such as dihydrotestosterone and testosterone) are AR ligands and act by binding to the AR in androgen-responsive tissues (Davey and Grossmann 2016). Human AR mutations and mouse knockout models have established a fundamental role for AR in masculinization and spermatogenesis (Maclean et al.; Walters et al. 2010; Rana et al. 2014). The AR is also expressed in many other tissues such as bone, muscles, ovaries and within the immune system (Rana et al. 2014).
Altered transcription of genes by the AR as a Key Event
Upon activation by ligand-binding, the AR translocates from the cytoplasm to the cell nucleus, dimerizes, binds to androgen response elements in the DNA to modulate gene transcription (Davey and Grossmann 2016). The transcriptional targets vary between cells and tissues, as well as with developmental stages and is also dependent on available co-regulators (Bevan and Parker 1999; Heemers and Tindall 2007). It should also be mentioned that the AR can work in other ‘non-canonial’ ways such as non-genomic signaling, and ligand-independent activation (Davey & Grossmann, 2016; Estrada et al, 2003; Jin et al, 2013).
A large number of known, and proposed, target genes of AR canonical signaling have been identified by analysis of gene expression following treatments with AR agonists (Bolton et al. 2007; Ngan et al. 2009, Jin et al. 2013).
How it is Measured or Detected
Altered transcription of genes by the AR can be measured by measuring the transcription level of known downstream target genes by RT-qPCR or other transcription analyses approaches, e.g. transcriptomics.
Since this KE aims to capture AR-mediated transcriptional patterns of effect, downstream bioinformatics analyses will typically be required to identify and compare effect footprints. Clusters of genes can be statistically associated with, for example, biological process terms or gene ontology terms relevant for AR-mediated signaling. Large transcriptomics data repositories can be used to compare transcriptional patterns between chemicals, tissues, and species (e.g. TOXsIgN (Darde et al, 2018a; Darde et al, 2018b), comparisons can be made to identified sets of AR ‘biomarker’ genes (e.g. as done in (Rooney et al, 2018)), and various methods can be used e.g. connectivity mapping (Keenan et al, 2019).
References
Bevan C, Parker M (1999) The role of coactivators in steroid hormone action. Exp. Cell Res. 253:349–356
Bolton EC, So AY, Chaivorapol C, et al (2007) Cell- and gene-specific regulation of primary target genes by the androgen receptor. Genes Dev 21:2005–2017. doi: 10.1101/gad.1564207
Darde, T. A., Gaudriault, P., Beranger, R., Lancien, C., Caillarec-Joly, A., Sallou, O., et al. (2018a). TOXsIgN: a cross-species repository for toxicogenomic signatures. Bioinformatics 34, 2116–2122. doi:10.1093/bioinformatics/bty040.
Darde, T. A., Chalmel, F., and Svingen, T. (2018b). Exploiting advances in transcriptomics to improve on human-relevant toxicology. Curr. Opin. Toxicol. 11–12, 43–50. doi:10.1016/j.cotox.2019.02.001.
Davey RA, Grossmann M (2016) Androgen Receptor Structure, Function and Biology: From Bench to Bedside. Clin Biochem Rev 37:3–15
Estrada M, Espinosa A, Müller M, Jaimovich E (2003) Testosterone Stimulates Intracellular Calcium Release and Mitogen-Activated Protein Kinases Via a G Protein-Coupled Receptor in Skeletal Muscle Cells. Endocrinology 144:3586–3597. doi: 10.1210/en.2002-0164
Heemers H V., Tindall DJ (2007) Androgen receptor (AR) coregulators: A diversity of functions converging on and regulating the AR transcriptional complex. Endocr. Rev. 28:778–808
Jin, Hong Jian, Jung Kim, and Jindan Yu. 2013. “Androgen Receptor Genomic Regulation.” Translational Andrology and Urology 2(3):158–77. doi: 10.3978/j.issn.2223-4683.2013.09.01
Keenan, A. B., Wojciechowicz, M. L., Wang, Z., Jagodnik, K. M., Jenkins, S. L., Lachmann, A., et al. (2019). Connectivity Mapping: Methods and Applications. Annu. Rev. Biomed. Data Sci. 2, 69–92. doi:10.1146/ANNUREV-BIODATASCI-072018-021211.
Maclean HE, Chu S, Warne GL, Zajact JD Related Individuals with Different Androgen Receptor Gene Deletions
MacLeod DJ, Sharpe RM, Welsh M, et al (2010) Androgen action in the masculinization programming window and development of male reproductive organs. In: International Journal of Andrology. Blackwell Publishing Ltd, pp 279–287
Ngan S, Stronach EA, Photiou A, et al (2009) Microarray coupled to quantitative RT–PCR analysis of androgen-regulated genes in human LNCaP prostate cancer cells. Oncogene 28:2051–2063. doi: 10.1038/onc.2009.68
Rana K, Davey RA, Zajac JD (2014) Human androgen deficiency: Insights gained from androgen receptor knockout mouse models. Asian J. Androl. 16:169–177
Rooney, J. P., Chorley, B., Kleinstreuer, N., and Corton, J. C. (2018). Identification of Androgen Receptor Modulators in a Prostate Cancer Cell Line Microarray Compendium. Toxicol. Sci. 166, 146–162. doi:10.1093/TOXSCI/KFY187.
Walters KA, Simanainen U, Handelsman DJ (2010) Molecular insights into androgen actions in male and female reproductive function from androgen receptor knockout models. Hum Reprod Update 16:543–558. doi: 10.1093/humupd/dmq003
List of Adverse Outcomes in this AOP
Event: 1786: Nipple retention (NR), increased
Short Name: nipple retention, increased
AOPs Including This Key Event
Biological Context
| Level of Biological Organization |
|---|
| Individual |
Domain of Applicability
Taxonomic Applicability Life Stage Applicability| Life Stage | Evidence |
|---|---|
| Birth to < 1 month | High |
| Sex | Evidence |
|---|---|
| Male | High |
The applicability domain of NR is limited to male laboratory strains of rats and mice from birth to juvenile age.
Key Event Description
In common laboratory strains of rats and mice, females typically have 6 (rats) or 5 (mice) pairs of nipples along the bilateral milk lines. In contrast, male rats and mice do not have nipples. This is unlike e.g., humans where both sexes have 2 nipples (Schwartz et al., 2021).
In laboratory rats, high levels of dihydrotestosterone (DHT) induce regression of the nipples in males (Imperato-McGinley & Gautier, 1986; Kratochwil, 1977; Kratochwil & Schwartz, 1976). Females, in the absence of this DHT surge, retain their nipples. This relationship has also been shown in numerous rat studies with perinatal exposure to anti-androgenic chemicals (Schwartz et al., 2021). Hence, if juvenile male rats and mice possess nipples, it is considered a sign of perturbed androgen action early in life.
This KE was first published by Pedersen et al (2022).
How it is Measured or Detected
Nipple retention (NR) is visually assessed, ideally on postnatal day (PND) 12/13 (OECD, 2018; Schwartz et al., 2021). However, PND 14 is also an accepted stage of examination (OECD, 2013). Depending on animal strain, the time when nipples become visible can vary, but the assessment of NR in males should be conducted when nipples are visible in their female littermates (OECD, 2013).
Nipples are detected as dark spots (or shadows) called areolae, which resemble precursors to a nipple rather than a fully developed nipple. The dark area may or may not display a nipple bud (Hass et al., 2007). Areolae typically emerge along the milk lines of the male pups corresponding to where female pups display nipples. Fur growth may challenge detection of areolae after PND 14/15. Therefore, the NR assessment should be conducted prior to excessive fur growth. Ideally, all pups in a study are assessed on the same postnatal day to minimize variation due to maturation level (OECD, 2013).
NR is occasionally observed in controls. Hence, accurate assessment of NR in controls is needed to detect substance-induced effects on masculine development (Schwartz et al., 2021). It is recommended by the OECD guidance documents 43 and 151 to record NR as a quantitative number rather than a qualitative measure (present/absent or yes/no response). This allows for more nuanced analysis of results, e.g., high control values may be recognized (OECD, 2013, 2018). Studies reporting quantitative measures of NR are therefore considered stronger in terms of weight of evidence.
Reproducibility of NR results is challenged by the measure being a visual assessment prone to a degree of subjectivity. Thus, NR should be assessed and scored blinded to exposure groups and ideally be performed by the same person(s) within the same study.
Regulatory Significance of the AO
NR is recognized by the OECD as a relevant measure for anti-androgenic effects and is mandatory in the test guidelines Extended One Generation Reproductive Toxicity Study, TG 443 (OECD, 2018) and the two screening studies for reproductive toxicity, TGs 421/422 (OECD, 2016a, 2016b). The endpoint is also described in the guidance documents 43 (OECD, 2008) and 151 (OECD, 2013). Furthermore, NR data can be used in chemical risk assessment for setting the No Observed Adverse Effect Level (NOAEL) as stated in the OECD guidance document 151 (OECD, 2013): “A statistically significant change in nipple retention should be evaluated similarly to an effect on AGD as both endpoints indicate an adverse effect of exposure and should be considered in setting a NOAEL”.
References
Hass, U., Scholze, M., Christiansen, S., Dalgaard, M., Vinggaard, A. M., Axelstad, M., Metzdorff, S. B., & Kortenkamp, A. (2007). Combined exposure to anti-androgens exacerbates disruption of sexual differentiation in the rat. Environmental Health Perspectives, 115(suppl 1), 122–128.
Imperato-McGinley, J., Binienda, Z., Gedney, J., & Vaughan, E. D. (1986). Nipple Differentiation in Fetal Male Rats Treated with an Inhibitor of the Enzyme 5α-Reductase: Definition of a Selective Role for Dihydrotestosterone. Endocrinology, 118(1), 132–137.
Kratochwil, K. (1977). Development and Loss of Androgen Responsiveness in the Embryonic Rudiment of the Mouse Mammary Gland. DEVELOPMENTAL BIOLOGY, 61, 358–365.
OECD. (2008). Guidance document 43 on mammalian reproductive toxicity testing and assessment. Environment, Health and Safety Publications, 16(43).
OECD. (2013). Guidance document supporting OECD test guideline 443 on the extended one-generation reproductive toxicity test. Environment, Health and Safety Publications, 10(151).
OECD. (2016a). Test Guideline 421: Reproduction/Developmental Toxicity Screening Test. OECD Guidelines for the Testing of Chemicals, 421. http://www.oecd.org/termsandconditions/
OECD. (2016b). Test Guideline 422: Combined Repeated Dose Toxicity Study with the Reproduction/Developmental Toxicity Screening Test. OECD Guidelines for the Testing of Chemicals, 422. http://www.oecd.org/termsandconditions/
OECD. (2018). Test Guideline 443: Extended one-generation reproductive toxicity study. OECD Guidelines for the Testing of Chemicals, 443. http://www.oecd.org/termsandconditions/
Pedersen, E. B., Christiansen, S., & Svingen, T. (2022). AOP key event relationship report: Linking androgen receptor antagonism with nipple retention. Current Research in Toxicology, 3, 100085.
Schwartz, C. L., Christiansen, S., Hass, U., Ramhøj, L., Axelstad, M., Löbl, N. M., & Svingen, T. (2021). On the Use and Interpretation of Areola/Nipple Retention as a Biomarker for Anti-androgenic Effects in Rat Toxicity Studies. Frontiers in Toxicology, 3, 730752.
Appendix 2
List of Key Event Relationships in the AOP
List of Adjacent Key Event Relationships
Relationship: 2130: Antagonism, Androgen receptor leads to Decrease, AR activation
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Androgen receptor (AR) antagonism leading to short anogenital distance (AGD) in male (mammalian) offspring | adjacent | High | High |
| Androgen receptor (AR) antagonism leading to nipple retention (NR) in male (mammalian) offspring | adjacent | High | |
| Androgen receptor (AR) antagonism leading to hypospadias in male (mammalian) offspring | adjacent | High | |
| Androgen receptor (AR) antagonism leading to decreased fertility in females | adjacent | High | Moderate |
Evidence Supporting Applicability of this Relationship
| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| mammals | mammals | High | NCBI |
| Life Stage | Evidence |
|---|---|
| During development and at adulthood | High |
| Sex | Evidence |
|---|---|
| Mixed | High |
This KER is applicable to mammals as AR expression and activity is highly conserved (Davey & Grossmann, 2016). AR activity is important for sexual development and reproduction in both males and females (Prizant et al., 2014; Walters et al., 2010). AR function is required during development, puberty, and adulthood. It is, however, acknowledged that this KER most likely has a much broader domain of applicability extending to non-mammalian vertebrates. AOP developers are encouraged to add additional relevant knowledge to expand on the applicability to also include other vertebrates.
Key Event Relationship Description
The androgen receptor (AR) is a ligand-activated steroid hormone nuclear receptor (Davey & Grossmann, 2016). In its inactive state, the AR locates to the cytoplasm (Roy et al., 2001). When activated, the AR translocates to the nucleus, dimerizes, and, together with co-regulators, binds to specific DNA regulatory sequences to regulate gene transcription (Davey & Grossmann, 2016) (Lamont and Tindall, 2010). This is considered the canonical AR signaling pathway. The AR can also activate non-genomic signalling (Jin et al., 2013). However, this KER focuses on the canonical pathway.
The two main AR ligands are the androgens testosterone (T) and the more potent dihydrotestosterone (DHT). Androgens bind to the AR to mediate downstream androgenic responses, such as male development and masculinization (Rey, 2021; Walters et al., 2010). Antagonism of the AR would decrease AR activation and therefore the downstream AR-mediated effects.
Evidence Supporting this KER
Biological PlausibilityThe biological plausibility for this KER is considered high.
The AR belongs to the steroid hormone nuclear receptor family. The AR has 3 main domains essential for its activity, the N-terminal domain, the ligand binding domain, and the DNA binding domain (Roy et al., 2001). Ligands, such as T and DHT, must bind to the ligand binding domain to activate AR allowing it to fulfill its role as a transcription factor. The binding of the ligand induces a change in AR conformation allowing it to translocate to the nucleus and congregate into a subnuclear compartment (Marcelli et al., 2006; Roy et al., 2001) homodimerize and bind to the DNA target sequences and regulate transcription of target genes. Regulation of AR target genes is greatly facilitated by numerous co-factors. Active AR signaling is essential for male reproduction and sexual development and is also crucial in several other tissues and organs such as ovaries, the immune system, bones, and muscles (Ogino et al., 2011; Prizant et al., 2014; Rey, 2021; William H. Walker, 2021).
AR antagonists can compete with or prevent in different ways AR ligand binding, thereby preventing AR activation. Antagonism of the AR can prevent translocation to the nucleus, compartmentalization, dimerization and DNA binding. Consequently, AR cannot regulate transcription of target genes and androgen signalling is disrupted. This can be observed using different AR activation assays such as AR dimerization, translocation, DNA binding or transcriptional activity assays (Brown et al., 2023; OECD, 2020).
Empirical EvidenceThe empirical evidence for this KER is considered high
The effects of AR antagonism have been shown in many studies in vivo and in vitro.
Several stressors can act as antagonists of the AR and lead to decreased AR activation. Some of these are detailed in an AOP key event relationship report by (Pedersen et al., 2022) and shown below, exhibiting evidence of dose-concordance:
Stressors
- Cyproterone acetate: Using the AR-CALUX reporter assay in antagonism mode, cyproterone acetate showed an IC50 of 7.1 nM (Sonneveld, 2005)
- Epoxiconazole: Using transiently AR-transfected CHO cells, epoxiconazole showed a LOEC of 1.6 µM and an IC50 of 10 µM (Kjærstad et al., 2010).
- Flutamide: Using the AR-CALUX reporter assay in antagonism mode, flutamide showed an IC50 of 1.3 µM (Sonneveld, 2005).
- Flusilazole: Using hAR-EcoScreen Assay, triticonazole showed a LOEC for antagonisms of 0.8 µM and an IC50 of 2.8 (±0.1) µM (Draskau et al., 2019).
- Prochloraz: Using transiently AR-transfected CHO cells, prochloraz showed a LOEC of 6.3 µM and an IC50 of 13 µM (Kjærstad et al., 2010).
- Propiconazole: Using transiently AR-transfected CHO cells, propiconazole showed a LOEC of 12.5 µM and an IC50 of 18 µM (Kjærstad et al., 2010).
- Tebuconazole: Using transiently AR-transfected CHO cells, tebuconazole showed a LOEC of 3.1 µM and an IC50 of 8.1 µM (Kjærstad et al., 2010).
- Triticonazole: Using hAR-EcoScreen Assay, triticonazole showed a LOEC for antagonisms of 0.2 µM and an IC50 of 0.3 (±0.01) µM (Draskau et al., 2019).
- Vinclozolin: Using the AR-CALUX reporter assay in antagonism mode, vinclozolin showed an IC50of 1.0 µM(Sonneveld, 2005).”(Pedersen et al., 2022)
Other evidence:
Known AR antagonists are used for treatment of AR-sensitive cancers such as flutamide for prostate cancer (Mahler et al., 1998).
Uncertainties and Inconsistencies
Known antiandrogenic compounds like hydroxyflutamide have been shown to act as agonists when the AR carries certain mutations, therefore contributing to uncertainties (Yeh et al., 1997). Additionally, the levels of endogenous androgens (e.g., testosterone or dihydrotestosterone) and the variability in the presence and function of AR co-activators may modulate the effect of AR antagonism.
Quantitative Understanding of the Linkage
Response-response relationshipThe quantitative relationship between AR antagonism and AR activation will depend on the type of antagonist.
Time-scaleNuclear translocation in HeLa cells transfected with AR-GFP show a response within 2 hours after ligand exposure (Marcelli et al., 2006; Szafran et al., 2008). Another assay focusing on AR binding to promoters in LNCaP cells has shown that after ligand binding, AR is able to translocate and bind to the DNA sequences within 15min showing the speed of AR activation (Kang et al., 2002).
Known Feedforward/Feedback loops influencing this KERAR antagonism can lead to increased AR transcript stability and levels as a compensatory mechanism in prostate cancer cells (Dart et al., 2020). In turn, in presence of increased AR levels, AR antagonists can exhibit agonistic activity (Chen et al., 2003).
References
Brown, E. C., Hallinger, D. R., Simmons, S. O., Puig-Castellví, F., Eilebrecht, E., Arnold, L., & Bioscience, P. A. (2023). High-throughput AR dimerization assay identifies androgen disrupting chemicals and metabolites. Front. Toxicol, 5, 1134783. https://doi.org/10.3389/ftox.2023.1134783
Chen, C. D., Welsbie, D. S., Tran, C., Baek, S. H., Chen, R., Vessella, R., Rosenfeld, M. G., & Sawyers, C. L. (2003). A R T I C L E S Molecular determinants of resistance to antiandrogen therapy. NATURE MEDICINE, 10(1). https://doi.org/10.1038/nm972
Dart, D. A., Ashelford, K., & Jiang, W. G. (2020). AR mRNA stability is increased with AR-antagonist resistance via 3′UTR variants. https://doi.org/10.1530/EC-19-0340
Davey, R. A., & Grossmann, M. (2016). Androgen Receptor Structure, Function and Biology: From Bench to Bedside. In Androgen Receptor Biology Clin Biochem Rev (Vol. 37, Issue 1).
Draskau, M. K., Boberg, J., Taxvig, C., Pedersen, M., Frandsen, H. L., Christiansen, S., & Svingen, T. (2019). In vitro and in vivo endocrine disrupting effects of the azole fungicides triticonazole and flusilazole. Environmental Pollution, 255, 113309. https://doi.org/10.1016/j.envpol.2019.113309
Jin, H. J., Kim, J., & Yu, J. (2013). Androgen receptor genomic regulation. In Translational Andrology and Urology (Vol. 2, Issue 3, pp. 158–177). AME Publishing Company. https://doi.org/10.3978/j.issn.2223-4683.2013.09.01
Kang, Z., Pirskanen, A., Jänne, O. A., & Palvimo, J. J. (2002). Involvement of Proteasome in the Dynamic Assembly of the Androgen Receptor Transcription Complex. Journal of Biological Chemistry, 277(50), 48366–48371. https://doi.org/10.1074/jbc.M209074200
Kjærstad, M. B., Taxvig, C., Nellemann, C., Vinggaard, A. M., & Andersen, H. R. (2010). Endocrine disrupting effects in vitro of conazole antifungals used as pesticides and pharmaceuticals. Reproductive Toxicology, 30(4), 573–582. https://doi.org/10.1016/j.reprotox.2010.07.009
Lamont, K. R., and Tindall, D. J. (2010). Androgen Regulation of Gene Expression. Adv. Cancer Res. 107, 137–162. doi:10.1016/S0065-230X(10)07005-3.
Mahler, C., Verhelst, J., and Denis, L. (1998). Clinical pharmacokinetics of the antiandrogens and their efficacy in prostate cancer. Clin. Pharmacokinet. 34, 405–417. doi:10.2165/00003088-199834050-00005/METRICS.
Marcelli, M., Stenoien, D. L., Szafran, A. T., Simeoni, S., Agoulnik, I. U., Weigel, N. L., Moran, T., Mikic, I., Price, J. H., & Mancini, M. A. (2006). Quantifying effects of ligands on androgen receptor nuclear translocation, intranuclear dynamics, and solubility. Journal of Cellular Biochemistry, 98(4), 770–788. https://doi.org/10.1002/jcb.20593
OECD (2020). Test No. 458: Stably Transfected Human Androgen Receptor Transcriptional Activation Assay for Detection of Androgenic Agonist and Antagonist Activity of Chemicals. OECD Guide. Paris: OECD Publishing doi:10.1787/9789264264366-en.
Ogino, Y., Miyagawa, S., Katoh, H., Prins, G. S., Iguchi, T., & Yamada, G. (2011). Essential functions of androgen signaling emerged through the developmental analysis of vertebrate sex characteristics. Evolution & Development, 13(3), 315–325. https://doi.org/10.1111/j.1525-142X.2011.00482.x
Pedersen, E. B., Christiansen, S., & Svingen, T. (2022). AOP key event relationship report: Linking androgen receptor antagonism with nipple retention. Current Research in Toxicology, 3, 100085. https://doi.org/10.1016/j.crtox.2022.100085
Prizant, H., Gleicher, N., & Sen, A. (2014). Androgen actions in the ovary: balance is key. Journal of Endocrinology, 222(3), R141–R151. https://doi.org/10.1530/JOE-14-0296
Rey, R. A. (2021). The Role of Androgen Signaling in Male Sexual Development at Puberty. Endocrinology, 162(2). https://doi.org/10.1210/endocr/bqaa215
Roy, A. K., Tyagi, R. K., Song, C. S., Lavrovsky, Y., Ahn, S. C., Oh, T. S., & Chatterjee, B. (2001). Androgen receptor: Structural domains and functional dynamics after ligand-receptor interaction. Annals of the New York Academy of Sciences, 949, 44–57. https://doi.org/10.1111/j.1749-6632.2001.tb04001.x
Sonneveld, E. (2005). Development of Androgen- and Estrogen-Responsive Bioassays, Members of a Panel of Human Cell Line-Based Highly Selective Steroid-Responsive Bioassays. Toxicological Sciences, 83(1), 136–148. https://doi.org/10.1093/toxsci/kfi005
Szafran, A. T., Szwarc, M., Marcelli, M., & Mancini, M. A. (2008). Androgen Receptor Functional Analyses by High Throughput Imaging: Determination of Ligand, Cell Cycle, and Mutation-Specific Effects. PLoS ONE, 3(11), e3605. https://doi.org/10.1371/journal.pone.0003605
Walters, K. A., Simanainen, U., & Handelsman, D. J. (2010). Molecular insights into androgen actions in male and female reproductive function from androgen receptor knockout models. In Human Reproduction Update (Vol. 16, Issue 5, pp. 543–558). Hum Reprod Update. https://doi.org/10.1093/humupd/dmq003
William H. Walker. (2021). Androgen Actions in the Testis and the Regulation of Spermatogenesis. In Advances in Experimental Medicine and Biology: Vol. volume 1381 (pp. 175–203).
Yeh, S., Miyamoto, H., & Chang, C. (1997). ARA70 and androgenic activity of hydroxyflutamide Hydroxyflutamide may not always be a pure antiandrogen (Vol. 349).
Relationship: 2124: Decrease, AR activation leads to Altered, Transcription of genes by the AR
AOPs Referencing Relationship
Evidence Supporting Applicability of this Relationship
| Term | Scientific Term | Evidence | Links |
|---|---|---|---|
| mammals | mammals | High | NCBI |
| Life Stage | Evidence |
|---|---|
| During development and at adulthood | High |
| Sex | Evidence |
|---|---|
| Mixed | High |
This KER is applicable for both sexes, across developmental stages into adulthood, in numerous cells and tissues and across mammalian taxa. It is, however, acknowledged that this KER most likely has a much broader domain of applicability extending to non-mammalian vertebrates. AOP developers are encouraged to add additional relevant knowledge to expand on the applicability to also include other vertebrates.
Key Event Relationship Description
The androgen receptor (AR) is a ligand-dependent nuclear transcription factor that upon activation translocates to the nucleus, dimerizes, and binds androgen response elements (AREs) to modulate transcription of target genes (Lamont and Tindall, 2010, Roy et al. 2001). Decreased activation of the AR affects its transcription factor activity, therefore leading to altered AR-target gene expression. This KER refers to decreased AR activation and altered gene expression occurring in complex systems, such as in vivo and the specific effect on transcription of AR target genes will depend on species, life stage, tissue, cell type etc.
Evidence Supporting this KER
Biological PlausibilityThe biological plausibility for this KER is considered high
The AR is a ligand-activated transcription factor part of the steroid hormone nuclear receptor family. Non-activated AR is found in the cytoplasm as a multiprotein complex with heat-shock proteins, immunophilins and, other chaperones (Roy et al. 2001). Upon activation through ligand binding, the AR dissociates from the protein complex, translocates to the nucleus and homodimerizes. Facilitated by co-regulators, AR can bind to DNA regions containing AREs and initiate transcription of target genes, that thus will be different in e.g. different tissues, life-stages, species etc.
Through mapping of AREs and ChIP sequencing studies, several AR target genes have been identified, mainly studied in prostate cells (Jin, Kim, and Yu 2013). Different co-regulators and ligands lead to altered expression of different sets of genes (Jin et al. 2013; Kanno et al. 2022). Alternative splicing of the AR can lead to different AR variants that also affects which genes are transcribed (Jin et al. 2013).
Apart from this canonical signaling pathway, the AR can suppress gene expression, indirectly regulate miRNA transcription, and have non-genomic effects by rapid activation of second messenger pathways in either presence or absence of a ligand (Jin et al. 2013).
Empirical EvidenceThe empirical evidence for this KER is considered high
In humans, altered gene expression profiling in individuals with androgen insensitivity syndrome (AIS) can provide supporting empirical evidence (Holterhus et al. 2003; Peng et al. 2021). In rodent AR knockout (KO) models, gene expression profiling studies and gene-targeted approaches have provided information on differentially expressed genes in several organ systems including male and female reproductive, endocrine, muscular, cardiovascular and nervous systems (Denolet et al. 2006; Fan et al. 2005; Holterhus et al. 2003; Ikeda et al. 2005; Karlsson et al. 2016; MacLean et al. 2008; Rana et al. 2011; Russell et al. 2012; Shiina et al. 2006; Wang et al. 2006; Welsh et al. 2012; Willems et al. 2010; Yu et al. 2008, 2012; Zhang et al. 2006; Zhou et al. 2011).
Exposure to known antiandrogens has been shown to alter transcriptional profiles, for example of neonatal pig ovaries (Knapczyk-Stwora et al. 2019).
Dose concordance has also been observed for instance in zebrafish embryos; a dose of 50 µg/L of the AR antagonist flutamide resulted in 674 differentially expressed genes at 96 h post fertilization whereas 500 µg/L flutamide resulted in 2871 differentially expressed genes (Ayobahan et al., 2023).
Uncertainties and InconsistenciesAR action has been reported to occur also without ligand binding. However, not much is known about the extent and biological implications of such non-canonical, ligand-independent AR activation (Bennesch and Picard 2015).
Quantitative Understanding of the Linkage
Response-response relationshipThere is not enough data to define a quantitative relationship between AR activation and alteration of AR target gene transcription, and such a relationship will differ between biological systems (species, tissue, cell type, life stage etc).
Time-scaleAR and promoter interactions occur within 15 minutes of ligand binding, RNA polymerase II and coactivator recruitment are proposed to occur transiently with cycles of approximately 90 minutes in LNCaP cells (Kang et al. 2002). RNA polymerase II elongation rates in mammalian cells have been shown to range between 1.3 and 4.3 kb/min (Maiuri et al. 2011). Therefore, depending on the cell type and the half-life of the AR target gene transcripts, changes are to be expected within hours.
Known modulating factors| Modulating Factor (MF) | MF Specification | Effect(s) on the KER | Reference(s) |
|---|---|---|---|
| Age | AR expression in aging male rats | Tissue-specific alterations in AR activity with aging | (Supakar et al. 1993; Wu, Lin, and Gore 2009) |
| Genotype | Number of CAG repeats in the first exon of AR | Decreased AR activation with increased number of CAGs |
(Tut et al. 1997; Chamberlain et al. 1994) |
AR has been hypothesized to auto-regulate its mRNA and protein levels (Mora and Mahesh 1999).
References
Ayobahan, S. U., Alvincz, J., Reinwald, H., Strompen, J., Salinas, G., Schäfers, C., et al. (2023). Comprehensive identification of gene expression fingerprints and biomarkers of sexual endocrine disruption in zebrafish embryo. Ecotoxicol. Environ. Saf. 250, 114514. doi:10.1016/J.ECOENV.2023.114514.
Bennesch, Marcela A., and Didier Picard. 2015. “Minireview: Tipping the Balance: Ligand-Independent Activation of Steroid Receptors.” Molecular Endocrinology 29(3):349–63.
Chamberlain, Nancy L., Erika D. Driverand, and Roger L. Miesfeldi. 1994. The Length and Location of CAG Trinucleotide Repeats in the Androgen Receptor N-Terminal Domain Affect Transactivation Function. Vol. 22.
Denolet, Evi, Karel De Gendt, Joke Allemeersch, Kristof Engelen, Kathleen Marchal, Paul Van Hummelen, Karen A. L. Tan, Richard M. Sharpe, Philippa T. K. Saunders, Johannes V. Swinnen, and Guido Verhoeven. 2006. “The Effect of a Sertoli Cell-Selective Knockout of the Androgen Receptor on Testicular Gene Expression in Prepubertal Mice.” Molecular Endocrinology 20(2):321–34. doi: 10.1210/me.2005-0113.
Fan, Wuqiang, Toshihiko Yanase, Masatoshi Nomura, Taijiro Okabe, Kiminobu Goto, Takashi Sato, Hirotaka Kawano, Shigeaki Kato, and Hajime Nawata. 2005. Androgen Receptor Null Male Mice Develop Late-Onset Obesity Caused by Decreased Energy Expenditure and Lipolytic Activity but Show Normal Insulin Sensitivity With High Adiponectin Secretion. Vol. 54.
Holterhus, Paul-Martin, Olaf Hiort, Janos Demeter, Patrick O. Brown, and James D. Brooks. 2003. Differential Gene-Expression Patterns in Genital Fibroblasts of Normal Males and 46,XY Females with Androgen Insensitivity Syndrome: Evidence for Early Programming Involving the Androgen Receptor. Vol. 4.
Ikeda, Yasumasa, Ken Ichi Aihara, Takashi Sato, Masashi Akaike, Masanori Yoshizumi, Yuki Suzaki, Yuki Izawa, Mitsunori Fujimura, Shunji Hashizume, Midori Kato, Shusuke Yagi, Toshiaki Tamaki, Hirotaka Kawano, Takahiro Matsumoto, Hiroyuki Azuma, Shigeaki Kato, and Toshio Matsumoto. 2005. “Androgen Receptor Gene Knockout Male Mice Exhibit Impaired Cardiac Growth and Exacerbation of Angiotensin II-Induced Cardiac Fibrosis.” Journal of Biological Chemistry 280(33):29661–66. doi: 10.1074/jbc.M411694200.
Jin, Hong Jian, Jung Kim, and Jindan Yu. 2013. “Androgen Receptor Genomic Regulation.” Translational Andrology and Urology 2(3):158–77.
Kang, Zhigang, Asta Pirskanen, Olli A. Jänne, and Jorma J. Palvimo. 2002. “Involvement of Proteasome in the Dynamic Assembly of the Androgen Receptor Transcription Complex.” Journal of Biological Chemistry 277(50):48366–71. doi: 10.1074/jbc.M209074200.
Kanno, Yuichiro, Nao Saito, Ryota Saito, Tomohiro Kosuge, Ryota Shizu, Tomofumi Yatsu, Takuomi Hosaka, Kiyomitsu Nemoto, Keisuke Kato, and Kouichi Yoshinari. 2022. “Differential DNA-Binding and Cofactor Recruitment Are Possible Determinants of the Synthetic Steroid YK11-Dependent Gene Expression by Androgen Receptor in Breast Cancer MDA-MB 453 Cells.” Experimental Cell Research 419(2). doi: 10.1016/j.yexcr.2022.113333.
Karlsson, Sara A., Erik Studer, Petronella Kettunen, and Lars Westberg. 2016. “Neural Androgen Receptors Modulate Gene Expression and Social Recognition but Not Social Investigation.” Frontiers in Behavioral Neuroscience 10(MAR). doi: 10.3389/fnbeh.2016.00041.
Knapczyk-Stwora, Katarzyna, Anna Nynca, Renata E. Ciereszko, Lukasz Paukszto, Jan P. Jastrzebski, Elzbieta Czaja, Patrycja Witek, Marek Koziorowski, and Maria Slomczynska. 2019. “Flutamide-Induced Alterations in Transcriptional Profiling of Neonatal Porcine Ovaries.” Journal of Animal Science and Biotechnology 10(1):1–15. doi: 10.1186/s40104-019-0340-y.
Lamont, K. R., and Tindall, D. J. (2010). Androgen Regulation of Gene Expression. Adv. Cancer Res. 107, 137–162. doi:10.1016/S0065-230X(10)07005-3.
MacLean, Helen E., W. S. Maria Chiu, Amanda J. Notini, Anna-Maree Axell, Rachel A. Davey, Julie F. McManus, Cathy Ma, David R. Plant, Gordon S. Lynch, and Jeffrey D. Zajac. 2008. “ Impaired Skeletal Muscle Development and Function in Male, but Not Female, Genomic Androgen Receptor Knockout Mice .” The FASEB Journal 22(8):2676–89. doi: 10.1096/fj.08-105726.
Maiuri, Paolo, Anna Knezevich, Alex De Marco, Davide Mazza, Anna Kula, Jim G. McNally, and Alessandro Marcello. 2011. “Fast Transcription Rates of RNA Polymerase II in Human Cells.” EMBO Reports 12(12):1280–85. doi: 10.1038/embor.2011.196.
Mora, Gloria R., and Virendra B. Mahesh. 1999. Autoregulation of the Androgen Receptor at the Translational Level: Testosterone Induces Accumulation of Androgen Receptor MRNA in the Rat Ventral Prostate Polyribosomes.
Peng, Yajie, Hui Zhu, Bing Han, Yue Xu, Xuemeng Liu, Huaidong Song, and Jie Qiao. 2021. “Identification of Potential Genes in Pathogenesis and Diagnostic Value Analysis of Partial Androgen Insensitivity Syndrome Using Bioinformatics Analysis.” Frontiers in Endocrinology 12. doi: 10.3389/fendo.2021.731107.
Rana, Kesha, Barbara C. Fam, Michele V Clarke, Tammy P. S. Pang, Jeffrey D. Zajac, and Helen E. Maclean. 2011. “Increased Adiposity in DNA Binding-Dependent Androgen Receptor Knockout Male Mice Associated with Decreased Voluntary Activity and Not Insulin Resistance.” Am J Physiol Endocrinol Me-Tab 301:767–78. doi: 10.1152/ajpendo.00584.2010.-In.
Roy, Arun K., Rakesh K. Tyagi, Chung S. Song, Yan Lavrovsky, Soon C. Ahn, Tae Sung Oh, and Bandana Chatterjee. 2001. “Androgen Receptor: Structural Domains and Functional Dynamics after Ligand-Receptor Interaction.” Pp. 44–57 in Annals of the New York Academy of Sciences. Vol. 949. New York Academy of Sciences.
Russell, Patricia K., Michele V. Clarke, Jarrod P. Skinner, Tammy P. S. Pang, Jeffrey D. Zajac, and Rachel A. Davey. 2012. “Identification of Gene Pathways Altered by Deletion of the Androgen Receptor Specifically in Mineralizing Osteoblasts and Osteocytes in Mice.” Journal of Molecular Endocrinology 49(1):1–10. doi: 10.1530/JME-12-0014.
Shiina, Hiroko, Takahiro Matsumoto, Takashi Sato, Katsuhide Igarashi, Junko Miyamoto, Sayuri Takemasa, Matomo Sakari, Ichiro Takada, Takashi Nakamura, Daniel Metzger, Pierre Chambon, Jun Kanno, Hiroyuki Yoshikawa, and Shigeaki Kato. 2006. Premature Ovarian Failure in Androgen Receptor-Deficient Mice. Vol. 103.
Supakar, P. C., C. S. Song, M. H. Jung, M. A. Slomczynska, J. M. Kim, R. L. Vellanoweth, B. Chatterjee, and A. K. Roy. 1993. “A Novel Regulatory Element Associated with Age-Dependent Expression of the Rat Androgen Receptor Gene.” Journal of Biological Chemistry 268(35):26400–408. doi: 10.1016/s0021-9258(19)74328-2.
Tut, Thein G., Farid J. Ghadessy, M. A. Trifiro, L. Pinsky, and E. L. Yong. 1997. Long Polyglutamine Tracts in the Androgen Receptor Are Associated with Reduced Trans-Activation, Impaired Sperm Production, and Male Infertility*. Vol. 82.
Wang, Ruey Sheng, Shuyuan Yeh, Lu Min Chen, Hung Yun Lin, Caixia Zhang, Jing Ni, Cheng Chia Wu, P. Anthony Di Sant’Agnese, Karen L. DeMesy-Bentley, Chii Ruey Tzeng, and Chawnshang Chang. 2006. “Androgen Receptor in Sertoli Cell Is Essential for Germ Cell Nursery and Junctional Complex Formation in Mouse Testes.” Endocrinology 147(12):5624–33. doi: 10.1210/en.2006-0138.
Welsh, M., L. Moffat, K. Belling, L. R. de França, T. M. Segatelli, P. T. K. Saunders, R. M. Sharpe, and L. B. Smith. 2012. “Androgen Receptor Signalling in Peritubular Myoid Cells Is Essential for Normal Differentiation and Function of Adult Leydig Cells.” International Journal of Andrology 35(1):25–40. doi: 10.1111/j.1365-2605.2011.01150.x.
Willems, Ariane, Sergio R. Batlouni, Arantza Esnal, Johannes V. Swinnen, Philippa T. K. Saunders, Richard M. Sharpe, Luiz R. França, Karel de Gendt, and Guido Verhoeven. 2010. “Selective Ablation of the Androgen Receptor in Mouse Sertoli Cells Affects Sertoli Cell Maturation, Barrier Formation and Cytoskeletal Development.” PLoS ONE 5(11). doi: 10.1371/journal.pone.0014168.
Wu, D. I., Grace Lin, and Andrea C. Gore. 2009. “Age-Related Changes in Hypothalamic Androgen Receptor and Estrogen Receptor in Male Rats.” The Journal of Comparative Neurology 512:688–701. doi: 10.1002/cne.21925.
Yu, I. Chen, Hung Yun Lin, Ning Chun Liu, Ruey Shen Wang, Janet D. Sparks, Shuyuan Yeh, and Chawnshang Chang. 2008. “Hyperleptinemia without Obesity in Male Mice Lacking Androgen Receptor in Adipose Tissue.” Endocrinology 149(5):2361–68. doi: 10.1210/en.2007-0516.
Yu, Shengqiang, Chiuan Ren Yeh, Yuanjie Niu, Hong Chiang Chang, Yu Chieh Tsai, Harold L. Moses, Chih Rong Shyr, Chawnshang Chang, and Shuyuan Yeh. 2012. “Altered Prostate Epithelial Development in Mice Lacking the Androgen Receptor in Stromal Fibroblasts.” Prostate 72(4):437–49. doi: 10.1002/pros.21445.
Zhang, Caixia, Shuyuan Yeh, Yen-Ta Chen, Cheng-Chia Wu, Kuang-Hsiang Chuang, Hung-Yun Lin, Ruey-Sheng Wang, Yu-Jia Chang, Chamindrani Mendis-Handagama, Liquan Hu, Henry Lardy, Chawnshang Chang, and † † George. 2006. Oligozoospermia with Normal Fertility in Male Mice Lacking the Androgen Receptor in Testis Peritubular Myoid Cells.
Zhou, Wei, Gensheng Wang, Christopher L. Small, Zhilin Liu, Connie C. Weng, Lizhong Yang, Michael D. Griswold, and Marvin L. Meistrich. 2011. “Erratum: Gene Expression Alterations by Conditional Knockout of Androgen Receptor in Adult Sertoli Cells of Utp14bjsd/Jsd (Jsd) Mice (Biology of Reproduction (2010) 83, (759-766) DOI: 10.1095/Biolreprod.110.085472).” Biology of Reproduction 84(2):400–408.
List of Non Adjacent Key Event Relationships
Relationship: 2133: Antagonism, Androgen receptor leads to nipple retention, increased
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| Androgen receptor (AR) antagonism leading to nipple retention (NR) in male (mammalian) offspring | non-adjacent | High |
Evidence Supporting Applicability of this Relationship
| Life Stage | Evidence |
|---|---|
| Foetal | High |
| Sex | Evidence |
|---|---|
| Male | High |
Taxonomic
The KER is considered directly applicable to rats and mice, in which males normally have no nipples due to high levels of androgens during development, leading to regression of nipple anlagen. The empirical evidence supports the relevance to rats, whereas the relevance in mice is assumed based on knowledge about developmental biology in this species. Applicability may extend to most rodents.
While NR is not directly translatable to humans, it serves as a clear indicator of diminished androgen activity causing disrupted fetal masculinisation and sexual differentiation during development - an effect considered relevant to mammals, humans (Schwartz et al., 2021) and vertebrates more broadly (Ogino et al., 2023). NR is included as a mandatory endpoint in several rodent OECD Test Guidelines (OECD 2025a; OECD 2025b, OECD 2025c) and in OECD GD 151 considered an adverse outcome applicable to the setting of Points of Departure for use in human health risk assessment (OECD, 2013). NR can also be used as an indicator of anti-androgenicity in mammals and vertebrates in the environment due to the conserved nature of the AR and its implication in sexual differentiation across species (Ogino et al., 2023).
Life stage
Programming of nipple/areola regression in males occurs during a short window of sensitivity to androgens in the nipple anlagen during fetal life. This takes place in rats around GD16-20 (Imperato-McGinley et al., 1986), which is, therefore, the relevant window of exposure. The relevant timing for the investigation of NR is PND12-14 in male rat offspring when the nipples are visible in the female littermates. At this time in development, the nipples/areolas are visible through the skin without excessive fur that may interfere with the investigation (Schwartz et al., 2021). It should be mentioned that though the occurrence of nipples/areolas in male offspring is believed to be relatively stable throughout life, it may be responsive to postnatal changes. Permanent nipple/areola retention is observed in some but not all in utero exposure studies with antiandrogens inducing nipple/areola retention at PND 12-14. Most of the differences between studies seem explainable by the window of exposure, dose levels and methods for investigation used, but the responsiveness of nipple/areola retention to postnatal changes remains to be fully explored (Schwartz et al., 2021).
Sex
Data presented in this KER support that disruption of androgen action during fetal life can lead to increased nipple/areola retention in male rat offspring. Since female mice and rat offspring, in general, have 10 (mice) or 12 (rats) nipples at the relevant time of investigation, increased nipple/areola retention at that time point is not a relevant endpoint for females.
Key Event Relationship Description
This KER links antagonism of the androgen receptor (AR) during fetal development to increased nipple/areola retention (NR) in male rodent offspring.
The KER is not directly applicable to humans, as both males and females have two nipples, and there is no known effect of androgens on their development (Schwartz et al., 2021). However, NR is a clear readout of reduced androgen action, fetal masculinization and sexual differentiation during development, which is relevant to humans, mammals (Schwartz et al., 2021), and vertebrates more broadly (Ogino et al., 2023). It is included as a mandatory endpoint in several rodent OECD Test Guidelines (OECD, 2025a, 2025b, 2025c) and, in OECD GD 151, is considered an adverse outcome applicable to the setting of Points of Departure for use in human health risk assessment (OECD, 2013). NR can also be used as an indicator of anti-androgenicity in mammals and vertebrates in the environment due to the conserved nature of the AR and its implication in sexual differentiation across species (Ogino et al., 2023).
Evidence Supporting this KER
Biological PlausibilityThe biological plausibility for this KER is judged to be high based on the following:
- Sexual differentiation happens in fetal life. The testes are developed and start to produce testosterone that is converted in other tissues by the enzyme 5-alpha-reductase to the more potent androgen dihydrotestosterone (DHT). Both hormones bind and activate the AR, which in turn drives the masculinization of the male fetus (Schwartz et al., 2021; Welsh et al., 2014).
- Fetal masculinization depends on the activation of androgen signalling during a critical time window, the masculinization programming window (MPW), from gestational day (GD) 16-20 in rats, 14.5-16.5 in mice and presumably gestation weeks (GWs) 8-14 in humans (Amato et al., 2022; Welsh et al., 2008).
- The fetal masculinization process involves a range of tissues and organs, including the nipple anlagen in rodents (primarily investigated in laboratory rats and mice). In humans, both sexes have two nipples. In contrast, rodents such as laboratory rats and mice are sexually dimorphic, with females having 12 (rats) and 10 (mice) nipples, and males generally having none (Mayer et al., 2008; Schwartz et al., 2021). In both male and female mouse embryos, stem cells differentiate into a mammary gland, with nipple anlagen being visible by embryonic day 11.5 (Mayer et al., 2008). In male embryos, the presence of androgen leads the nipple anlagen to regress a few days later (Kratochwil, 1977; Kratochwil & Schwartz, 1976) . The androgen responsiveness in the nipple anlagen is rather short, in mice, starting late embryonic day 13, with loss of responsiveness on embryonic day 15 (Imperato-McGinley et al., 1986; Kratochwil, 1977) and thus roughly following the timing of the MPW.
- Nipple formation is inhibited in female mouse and rat fetuses exposed to androgens during gestation (Goldman et al., 1976; Greene et al., 1941; Imperato-McGinley et al., 1986).
- Male Tfm-mutant mice, which are insensitive to androgens and believed to be so due to a nonfunctional AR, present with retained nipples (Kratochwil & Schwartz, 1976).
- Multiple mechanisms of action may potentially lead to NR in male mouse and rat offspring. DHT is the main androgen responsible for regression of the nipples through interaction with AR in the nipple anlagen (Imperato-McGinley et al., 1986). Inhibition of testosterone synthesis or the conversion of testosterone to DHT, increased metabolism of androgens, or direct interference with AR activation may thus all lead to nipple/areola retention (Imperato-McGinley et al., 1986; Schwartz et al., 2021).
Empirical Evidence
The empirical support from studies in animals for this KER is judged as high overall.
The relationship is supported by numerous studies demonstrating induction of NR in male offspring after in utero exposure to substances known to antagonise AR in vitro (fenitrothion, flutamide, linuron, mancozeb, pp’-DDE, prochloraz, procymidone, pyrifluquinazon, tebuconazole and vinclozolin) (Pedersen et al., 2022; Appendix 1, 12h4h48cc2_KER_2133_Appendix_1.pdf). The empirical evidence includes only studies conducted in rats, although it is believed that the link also exists in mice, and other rodent species (Pedersen et al., 2022). Some inconsistences in the empirical evidence for 3 of the substances were observed. These could, however, be explained by differences in dose levels, and the level of confidence for prenatal exposure to these substances resulting in increased NR in male offspring is judged to be strong (Appendix 1,12h4h48cc2_KER_2133_Appendix_1.pdf).
Dose concordance
Dose concordance is challenging to assess for this KER since the upstream event is measured in vitro and the downstream event is measured in vivo.
Temporal concordance
Temporal concordance can only be considered from a theoretical perspective since the downstream event, ‘increased NR’, is a result of the disruption of the normal regression of nipple anlagen in male rodents induced during a short window of gestational development (in mice of approximately 2 days), but usually measured at PND12-14 in rats. Earlier than this, the areolae are not yet visible through the skin and later than this, the animals grow fur and need to be shaved for proper examination. This is supported by several of the studies in the empirical evidence, where the test substance was administered during a short period during gestation and nipple retention was observed postnatally.
Based on current knowledge, it is understood that the upstream event – antagonism of the AR – takes place minutes to hours after exposure to an anti-androgenic substance.
Uncertainties and InconsistenciesA major challenge with using NR as a biomarker is the subjectivity of the measurement. In juvenile rat pups, nipples are only present as areolae, i.e., dark shadows with or without a nipple bud. This means that the experience of the personnel assessing the presence and number of areolae/nipples can influence the results. Furthermore, the results are likely prone to larger variation if several assessors are used to record NR within the same study. To minimise these sources of uncertainty, assessors must be trained to recognise areolae and not look for fully developed nipples. Moreover, the number of assessors should be limited to one or two, and they should always be blinded to exposure groups.
Another factor that may affect NR results is the age of the rat pups at the time of assessment. OECD guidelines have standardised the time for measuring the occurrence of NR to be optimal at PD 12 or 13, when they are visible in female littermates (OECD, 2013).
Quantitative Understanding of the Linkage
The quantitative understanding of the relationship between decreased AR activity and NR is challenged by the fact that the potency of AR antagonism in vitro is not proportional to the magnitude of NR observed in vivo (Gray et al., 2019).
Response-response relationshipThe difficulties in extrapolating potency from in vitro to in vivo studies were exemplified by a comparison of the effects of pyrifluquinazon and bisphenol C in vitro and in utero. In vitro, bisphenol C antagonized the androgen receptor with a much higher potency than pyrifluquinazon, but in vivo the potencies were reversed with pyrifluquinazon exposure leading to NR at lower exposure levels than bisphenol C (Gray et al., 2019).
Time-scaleAR activation operates on a time-scale of minutes. The AR is a ligand-activated nuclear receptor and transcription factor. Upon ligand binding a conformational change and subsequent dimerisation of the AR take place within 3-6 minutes (Schaufele et al., 2005). Nuclear translocation (Nightingale et al., 2003) and promoter interactions occur within 15 minutes of ligand binding, and RNA polymerase II and coactivator recruitment are then proposed to occur transiently with cycles of approximately 90 minutes (Kang et al., 2002).
For the downstream event, the time-scale for observing a measurable effect on nipple/areola retention is closer to days and weeks, depending on species. For instance, in mice, the nipple anlage are responsive to androgen action at embryonic day 13-15, while a sexual dimorphism of the nipples/areolas can first be observed after birth (Imperato-McGinley et al., 1986) .
Known modulating factorsA well-established modulating factor for androgen action is genetic variations in the AR, which decrease the function of the receptor. For example, longer CAG repeat lengths have been associated with decreased AR activation (Chamberlain et al., 1994; Tut et al., 1997).
Rat strain is another important modulating factor, with studies showing that the Long-Evans Hooded rat is less sensitive to nipple/areola retention than the Sprague-Dawley rat (Wolf et al., 1999; You et al., 1998).
Known Feedforward/Feedback loops influencing this KERNot relevant for this KER.
References
Amato, C. M., Yao, H. H.-C., & Zhao, F. (2022). One Tool for Many Jobs: Divergent and Conserved Actions of Androgen Signaling in Male Internal Reproductive Tract and External Genitalia. Frontiers in Endocrinology, 13. https://doi.org/10.3389/fendo.2022.910964
Chamberlain, N. L., Driver, E. D., & Miesfeld, R. L. (1994). The length and location of CAG trinucleotide repeats in the androgen receptor N-terminal domain affect transactivation function. Nucleic Acids Research, 22(15), 3181–3186. https://doi.org/10.1093/nar/22.15.3181
Goldman AS, Shapiro B, & Neumann F. (1976). Role of testosterone and its metabolites in the differentiation of the mammary gland in rats. Endocrinology, 99(6), 1490–1495. https://doi.org/10.1210/endo-99-6-1490
Gray, L. E., Furr, J. R., Conley, J. M., Lambright, C. S., Evans, N., Cardon, M. C., Wilson, V. S., Foster, P. M., & Hartig, P. C. (2019). A Conflicted Tale of Two Novel AR Antagonists in Vitro and in Vivo: Pyrifluquinazon Versus Bisphenol C. Toxicological Sciences, 168(2), 632–643. https://doi.org/10.1093/toxsci/kfz010
Greene, R. R., Burrill, M. W., & Ivy, A. C. (1941). Experimental intersexuality: The effects of combined estrogens and androgens on the embryonic sexual development of the rat. Journal of Experimental Zoology, 87(2), 211–232. https://doi.org/10.1002/jez.1400870203
Imperato-McGinley J, Binienda Z, Gedney J, & Vaughan ED Jr. (1986). Nipple differentiation in fetal male rats treated with an inhibitor of the enzyme 5 alpha-reductase: definition of a selective role for dihydrotestosterone. Endocrinology, 118(1), 132–137. https://doi.org/10.1210/endo-118-1-132
Kang, H.-Y., Huang, K.-E., Chang, S. Y., Ma, W.-L., Lin, W.-J., & Chang, C. (2002). Differential Modulation of Androgen Receptor-mediated Transactivation by Smad3 and Tumor Suppressor Smad4. Journal of Biological Chemistry, 277(46), 43749–43756. https://doi.org/10.1074/jbc.M205603200
Kratochwil, K. (1977). Development and loss of androgen responsiveness in the embryonic rudiment of the mouse mammary gland. Developmental Biology, 61(2), 358–365. https://doi.org/10.1016/0012-1606(77)90305-0
Kratochwil, K., & Schwartz, P. (1976). Tissue interaction in androgen response of embryonic mammary rudiment of mouse: identification of target tissue for testosterone. Proceedings of the National Academy of Sciences, 73(11), 4041–4044. https://doi.org/10.1073/pnas.73.11.4041
Mayer, J. A., Foley, J., De La Cruz, D., Chuong, C. M., & Widelitz, R. (2008). Conversion of the nipple to hair-bearing epithelia by lowering bone morphogenetic protein pathway activity at the dermal-epidermal interface. The American Journal of Pathology, 173(5), 1339–1348. https://doi.org/10.2353/AJPATH.2008.070920
Nightingale, J., Chaudhary, K. S., Abel, P. D., Stubbs, A. P., Romanska, H. M., Mitchell, S. E., Stamp, G. W. H., & Lalani, E.-N. (2003). Ligand Activation of the Androgen Receptor Downregulates E-Cadherin-Mediated Cell Adhesion and Promotes Apoptosis of Prostatic Cancer Cells. Neoplasia, 5(4), 347–361. https://doi.org/10.1016/S1476-5586(03)80028-3
OECD (2013), Guidance Document Supporting OECD Test Guideline 443 on the Extended One-Generational Reproductive Toxicity Test, OECD Series on Testing and Assessment, No. 151, OECD Publishing, Paris, ENV/JM/MONO(2013)10
OECD (2025a), Test No. 443: Extended One-Generation Reproductive Toxicity Study, OECD Guidelines for the Testing of Chemicals, Section 4, OECD Publishing, Paris, https://doi.org/10.1787/9789264185371-en.
OECD (2025b), Test No. 421: Reproduction/Developmental Toxicity Screening Test, OECD Guidelines for the Testing of Chemicals, Section 4, OECD Publishing, Paris, https://doi.org/10.1787/9789264264380-en.
OECD (2025c), Test No. 422: Combined Repeated Dose Toxicity Study with the Reproduction/Developmental Toxicity Screening Test, OECD Guidelines for the Testing of Chemicals, Section 4, OECD Publishing, Paris, https://doi.org/10.1787/9789264264403-en.
Ogino, Y., Ansai, S., Watanabe, E. et al. Evolutionary differentiation of androgen receptor is responsible for sexual characteristic development in a teleost fish. Nat Commun 14, 1428 (2023). https://doi.org/10.1038/s41467-023-37026-6
Pedersen, E. B., Christiansen, S., & Svingen, T. (2022). AOP key event relationship report: Linking androgen receptor antagonism with nipple retention. In Current Research in Toxicology (Vol. 3). Elsevier B.V. https://doi.org/10.1016/j.crtox.2022.100085
Schwartz, C. L., Christiansen, S., Hass, U., Ramhøj, L., Axelstad, M., Löbl, N. M., & Svingen, T. (2021). On the Use and Interpretation of Areola/Nipple Retention as a Biomarker for Anti-androgenic Effects in Rat Toxicity Studies. In Frontiers in Toxicology (Vol. 3). Frontiers Media S.A. https://doi.org/10.3389/ftox.2021.730752
Tut, T. G., Ghadessy, F. J., Trifiro, M. A., Pinsky, L., & Yong, E. L. (1997). Long Polyglutamine Tracts in the Androgen Receptor Are Associated with Reduced Trans -Activation, Impaired Sperm Production, and Male Infertility 1. The Journal of Clinical Endocrinology & Metabolism, 82(11), 3777–3782. https://doi.org/10.1210/jcem.82.11.4385
Welsh, M., Saunders, P. T. K., Fisken, M., Scott, H. M., Hutchison, G. R., Smith, L. B., & Sharpe, R. M. (2008). Identification in rats of a programming window for reproductive tract masculinization, disruption of which leads to hypospadias and cryptorchidism. JOURNAL OF CLINICAL INVESTIGATION, 118(4), 1479–1490. https://doi.org/10.1172/JCI34241
Welsh, M., Suzuki, H., & Yamada, G. (2014). The Masculinization Programming Window. In O. Hiort & S. F. Ahmed (Eds.), UNDERSTANDING DIFFERENCES AND DISORDERS OF SEX DEVELOPMENT (DSD) (Vol. 27, pp. 17–27). https://doi.org/10.1159/000363609
Wolf C Jr, Lambright C, Mann P, Price M, Cooper RL, Ostby J, Gray LE Jr. Administration of potentially antiandrogenic pesticides (procymidone, linuron, iprodione, chlozolinate, p,p'-DDE, and ketoconazole) and toxic substances (dibutyl- and diethylhexyl phthalate, PCB 169, and ethane dimethane sulphonate) during sexual differentiation produces diverse profiles of reproductive malformations in the male rat. Toxicol Ind Health. 1999 Jan-Mar;15(1-2):94-118. doi: 10.1177/074823379901500109. PMID: 10188194.
You, L., Casanova, M., Archibeque-Engle, S., Sar, M., Fan, L.-Q., & Heck, A. (1998). Impaired Male Sexual Development in Perinatal Sprague-Dawley and Long-Evans Hooded Rats Exposed in Utero and Lactationally to p,p’-DDE. In TOX1COLOGICAL SCIENCES (Vol. 45). https://academic.oup.com/toxsci/article/45/2/162/1653877
Relationship: 3348: Decrease, AR activation leads to nipple retention, increased
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding |
|---|---|---|---|
| 5α-reductase inhibition leading to increased nipple retention (NR) in male (rodent) offspring | non-adjacent | High | |
| Androgen receptor (AR) antagonism leading to nipple retention (NR) in male (mammalian) offspring | non-adjacent | High | |
| Decreased testosterone synthesis leading to increased nipple retention (NR) in male (rodent) offspring | non-adjacent | High |
Evidence Supporting Applicability of this Relationship
| Life Stage | Evidence |
|---|---|
| Foetal | High |
| Sex | Evidence |
|---|---|
| Male | High |
The KER is considered directly applicable to rats and mice, in which males normally have no nipples due to high levels of androgens during development, leading to regression of nipple anlagen. The empirical evidence supports the relevance to rats, whereas the relevance in mice is assumed based on knowledge about developmental biology in this species. Applicability may extend to most rodents.
While NR is not directly translatable to humans, it serves as a clear indicator of diminished androgen activity causing disrupted fetal masculinisation and sexual differentiation during development - an effect considered relevant to mammals, humans (Schwartz et al., 2021), and vertebrates more broadly (Ogino et al., 2023). NR is included as a mandatory endpoint in several rodent OECD Test Guidelines (OECD 2025a; OECD 2025b, OECD 2025c) and in OECD GD 151 considered an adverse outcome applicable to the setting of Points of Departure for use in human health risk assessment (OECD, 2013). NR can also be used as an indicator of anti-androgenicity in mammals and vertebrates in the environment due to the conserved nature of the AR and its implication in sexual differentiation across species (Ogino et al., 2023).
Life stage
Programming of nipple/areola regression in males occurs during a short window of sensitivity to androgens in the nipple anlagen during fetal life. This takes place in rats around embryonic days 13-15 (Imperato-McGinley et al., 1986), which is, therefore, the relevant window of exposure. The relevant timing for the investigation of NR is PND12-14 in male rat offspring when the nipples are visible in the female littermates. At this time in development, the nipples/areolas are visible through the skin without excessive fur that may interfere with the investigation (Schwartz et al., 2021). It should be mentioned that though the occurrence of nipples/areolas in male offspring is believed to be relatively stable throughout life, it may be responsive to postnatal changes. Permanent nipple/areola retention is observed in some but not all in utero exposure studies with antiandrogens inducing nipple/areola retention at PND 12-14. Most of the differences between studies seem explainable by the window of exposure, dose levels and methods for investigation used, but the responsiveness of nipple/areola retention to postnatal changes remains to be fully explored (Schwartz et al., 2021).
Sex
Data presented in this KER support that disruption of androgen action during fetal life can lead to increased nipple/areola retention in male rat offspring. Since female mice and rat offspring, in general, have 10 (mice) or 12 (rats) nipples at the relevant time of investigation, increased nipple/areola retention at that time point is not a relevant endpoint for females.
Key Event Relationship Description
This KER links a decrease in androgen receptor (AR) activation during fetal development to increased nipple/areola retention (NR) in male rodent offspring. It should be noted that the upstream Key Event (KE) ‘decrease, androgen receptor activation’ (KE-1614 in AOP Wiki) specifically focuses on decreased activation of the AR in vivo, while most methods that can be used to measure AR activity are carried out in vitro. Indirect information about this KE may, for example, be provided from assays showing in vitro AR antagonism, decreased in vitro or in vivo testosterone production/levels, or decreased in vitro or in vivo dihydrotestosterone (DHT) production/levels.
The KER is not directly applicable to humans as both sexes have two nipples, and there is no known effect of androgens on their development (Schwartz et al., 2021). However, NR is a clear readout of reduced androgen action, fetal masculinization and sexual differentiation during development, which is relevant to humans, mammals (Schwartz et al., 2021), and vertebrates more broadly (Ogino et al., 2023). It is included as a mandatory endpoint in several rodent OECD Test Guidelines (OECD, 2025a, 2025b, 2025c) and, in OECD GD 151, is considered an adverse outcome applicable to the setting of Points of Departure for use in human health risk assessment (OECD, 2013). NR can also be used as an indicator of anti-androgenicity in mammals and vertebrates in the environment due to the conserved nature of the AR and its implication in sexual differentiation across species (Ogino et al., 2023).
Evidence Supporting this KER
Biological PlausibilityThe biological plausibility for this KER is judged to be high based on the following:
- Sexual differentiation happens in fetal life. The testes are developed and start to produce testosterone that is converted in other tissues by the enzyme 5-alpha-reductase to the more potent androgen dihydrotestosterone (DHT). Both hormones bind and activate the nuclear receptor and transcription factor AR, which in turn drives the masculinization of the male fetus (Schwartz et al., 2021; Welsh et al., 2014).
- Fetal masculinization depends on the activation of androgen signalling during a critical time window, the masculinization programming window (MPW), from gestational day (GD) 16-20 in rats, 14.5-16.5 in mice and presumably gestation weeks (GWs) 8-14 in humans (Amato et al., 2022; Welsh et al., 2008).
- The fetal masculinization process involves a range of tissues and organs, including the nipple anlagen in rats and mice. In humans, both sexes have two nipples. In contrast, common laboratory mice and rats are sexually dimorphic, with females having 12 (rats) and 10 (mice) nipples and males generally having none (Mayer et al., 2008; Schwartz et al., 2021). In both male and female mouse embryos, stem cells differentiate into a mammary gland, with nipple anlagen being visible by embryonic day 11.5 (Mayer et al., 2008). In male embryos, the presence of androgen leads the nipple anlagen to regress a few days later (Kratochwil, 1977; Kratochwil & Schwartz, 1976) . The androgen responsiveness in the nipple anlagen is rather short, in mice starting late embryonic day 13, with loss of responsiveness on embryonic day 15 (Imperato-McGinley et al., 1986; Kratochwil, 1977) and thus roughly following the timing of the MPW.
- Nipple formation is inhibited in female mice and rat fetuses exposed to androgens during gestation (Goldman et al., 1976; Greene et al., 1941; Imperato-McGinley et al., 1986).
- Male Tfm-mutant mice, which are insensitive to androgens and believed to be so due to a nonfunctional androgen receptor, present with retained nipples (Kratochwil & Schwartz, 1976).
- Multiple mechanisms of action may potentially lead to nipple retention in male mouse and rat offspring. DHT is the main androgen responsible for nipple/areola regression through interaction with AR in the nipple anlagen (Imperato-McGinley et al., 1986). Inhibition of testosterone synthesis or the conversion of testosterone to DHT, increased metabolism of androgens, or direct interference with AR activation may thus all lead to nipple/areola retention (Imperato-McGinley et al., 1986; Schwartz et al., 2021).
Empirical EvidenceThe empirical support from studies in animals for this KER is judged as high overall.
It should be noted that the KE decreased AR activation (KE 1614 in AOP Wiki) specifically focuses on decreased activation of the AR in vivo, with no methods currently available to measure this. Examples of assays that provide indirect information about KE 1614 are described in upstream MIE/KEs.
The empirical evidence for this KER from animal studies in vivo is based on studies using six different substances that result in decreased AR activation by different mechanisms. Flutamide, procymidone and vinclozolin bind to the AR and inhibit the receptor activity and thereby act as AR antagonists, see MIE 26. Finasteride inhibits the 5-alpha-reductase enzyme that converts testosterone to DHT, see MIE 1617. DEHP and DBP exposure during prenatal development in rats results in reduced fetal testosterone levels, see KE-2298 and KE1690. (MIE 26, MIE 1617 and KE 1690 can be found in AOP-Wiki).
The evidence for the upstream KE is mainly based on data from in vitro assays (AR antagonism or 5-alpha-reductase inhibition in vitro), whereas the evidence for the downstream KE is based on in vivo studies, and there is generally no evidence for both KEs from the same study. However, decreased testosterone levels can be measured in vivo, and (Howdeshell et al., 2007; Martino-Andrade et al., 2009) measured the effect of developmental phthalate exposure on both testosterone levels and nipple/areola retention (see the section about “Dose concordance”).
The empirical evidence for the six substances is summarised in Table 3.
Table 3. Summary of empirical evidence for decreased androgen receptor activation, leading to decreased nipple/areola retention. References for the studies supporting the empirical evidence are found in the section “Evidence for decreased AR activation (KE 1614) by flutamide, procymidone, and vinclozolin, finasteride, DEHP and DBP” and in Table 4 in Appendix 2 (6djoma9gmj_KER_3348_Appendix_2_.pdf).
|
Stressor(s) |
Upstream effect |
Downstream effect |
|
Flutamide |
AR antagonism in in vitro assay, receptor binding and transactivation assays
|
Increased NR in males after prenatal exposure in studies in rat
|
|
Procymidone |
AR antagonism in in vitro assay receptor binding and transactivation assays
|
Increased NR in males after prenatal exposure in studies in rat
|
|
Vinclozolin |
AR antagonism in in vitro assay receptor binding and transactivation assays
|
Increased NR in males after prenatal exposure in studies in rat |
|
Finasteride |
Inhibition of 5-alpha-reductase enzyme in in vitro assays
|
Increased NR in males after prenatal exposure in studies in rat
|
|
DEHP |
Reduced production of testosterone in fetal testis measured in ex vivo testis assays, reduced testosterone levels in testis, and reduced fetal plasma or serum testosterone levels
|
Increased NR in males after prenatal exposure in studies in rat
|
|
DBP |
Reduced production of testosterone in fetal testis measured in ex vivo testis assays and reduced testosterone levels in fetal testis |
Increased NR in males after prenatal exposure in studies in rat
|
From Table 3, it can be deduced that fetal exposure to substances known to decrease androgen receptor activation through antagonism of the AR (vinclozolin, procymidone, flutamide), inhibition of testosterone synthesis (DEHP, DBP) or inhibition of the conversion of testosterone to DHT (finasteride), results in increased nipple/areola retention in rat male offspring.
Evidence for decreased AR activation (KE 1614) by flutamide, procymidone, vinclozolin, finasteride, DEHP and DBP.
Flutamide, a pharmaceutical, binds the AR and inhibits its activity, thereby acting as an AR antagonist. It has been used as an antiandrogen for the treatment of prostate cancer and is used as a reference chemical for antiandrogenic activity in the AR transactivation assays in the OECD test guideline No 458 (Goldspiel & Kohler, 1990; Labrie, 1993; OECD, 2023; Simard et al., 1986)
Procymidone and vinclozolin are fungicides that have been shown to be AR antagonists. Procymidone binds to the AR and inhibits the agonist binding, as shown in AR binding assays using rat prostate cytosol (Hosokawa et al., 1993) or AR transfected cells (Ostby et al., 1999). Procymidone also inhibits agonist activated transcription in AR reporter assays (Hass et al., 2012; Kojima et al., 2004; Orton et al., 2011; Ostby et al., 1999; Scholze et al., 2020). Vinclozolin binds to the AR and inhibits the agonist binding, as shown in AR binding assays using rat epididymis cytosol (Kelce & Wilson, 1997) or AR transfected cells (Wong et al., 1995). Vinclozolin also inhibits agonist activated transcription in AR reporter assays (Euling, 2002; Kojima et al., 2004; Molina-Molina et al., 2006; Orton et al., 2011; Scholze et al., 2020; Shimamura et al., 2002; Wong et al., 1995).
Finasteride is a pharmaceutical that inhibits the 5-alpha-reductase enzyme that converts testosterone to DHT. Finasteride is used to treat benign prostatic hypertrophy (Andersson & Russell, 1990; Stoner, 1990; Wood & Rittmaster, 1994).
Prenatal exposure to DEHP in rats has been shown to reduce the production of testosterone in fetal testis measured in ex vivo testis assays, and to reduce testosterone levels in testis and in fetal plasma and serum (Borch et al., 2006; Borch J et al., 2004; Culty et al., 2008; Hannas et al., 2011, 2012; Howdeshell et al., 2007; Klinefelter et al., 2012; Parks, 2000; VO et al., 2009; Wilson et al., 2004, 2007). Conversely, prenatal DEHP exposure did not result in any effects on testosterone levels in the testis at PND1 in one study by Andrade et al. (2006) (Andrade et al., 2006). Similar to DEHP, prenatal exposure to DBP has been shown to reduce the production of testosterone in fetal rat testis measured in ex vivo testis studies (Howdeshell et al., 2007; Wilson et al., 2004) and reduce testosterone levels in the fetal rat testis (Martino-Andrade et al., 2009). The precise underlying mechanism for these effects of DEHP and DPB is presently unknown.
Evidence for increased nipple/areola retention in males (AO-1786) by prenatal exposure to flutamide, procymidone, vinclozolin, finasteride, DEHP and DBP.
All datasets that were used for the weight of evidence assessment were judged as reliable without or with restriction. The majority of datasets assessed showed an increased nipple/areola retention in male offspring after gestational exposure. The conclusion was that the level of confidence was strong for all six substances. The studies are summarised in Table 4 in Appendix 2, 6djoma9gmj_KER_3348_Appendix_2_.pdf
Dose concordance
Dose concordance is challenging to assess for this KER since in vivo AR activity is currently not possible to measure, but can only be inferred indirectly by measures of upstream events. In some studies, fetal (testicular) testosterone levels during, or close to, the fetal masculinization programming window are measured in a subset of animals exposed similarly to those investigated for NR post-natally. Such information may inform on dose concordance if more doses are included.
In a rat in utero exposure study (GD13-21) with DPB and DEHP, testosterone levels in the fetal testes were investigated at GD21, and NR was investigated at PND13 (Martino-Andrade et al., 2009). For DBP, both reduced testosterone levels in fetal testes and NR were observed at 500 mg/kg/d, whereas no effect on NR and only a slight non-significant reduction of testosterone was observed at the lower dose (100 mg/kg/d). For DEHP, a slight but non-significant decrease in testosterone levels in fetal rat testis was observed after exposure to 150 mg/kg/d DEHP, with no effects on nipple/areola retention.
Such data could suggest dose concordance for this part of the KER, although the evidence for this is not strong.
Temporal concordance
Temporal concordance can only be considered from a theoretical perspective since the downstream event, increased NR, is a result of disruption to normal regression of nipple anlagen in male rodents induced during a short window of gestational development (in mice of approximately 2 days), but usually measured at PND12-14 in rats. Earlier than this, the areolae are not yet visible through the skin and later than this, the animals grow fur and need to be shaved for proper examination. This is supported by several of the studies in the empirical evidence, where the test substance was administered during a short period during gestation and nipple retention was observed postnatally.
Based on current knowledge, it is understood that the upstream event – decreased AR activation in vivo – takes place minutes to hours after exposure to an anti-androgenic substance. If a substance decreases AR activation through inhibition of the AR, the upstream event is expected to happen immediately after exposure. If a substance decreases androgen receptor activation through inhibition of testosterone synthesis, the upstream event is expected to happen minutes to hours after the exposure.
Uncertainties and Inconsistencies
For DEHP and DBP, there were some inconsistencies in the empirical evidence, but they could be explained by differences in study designs and uncertainties in measurements (see Appendix 1). Some uncertainty is imposed by the poorly supported dose-concordance. However, the dose-concordance is well supported by the current understanding of biological processes.
Quantitative Understanding of the Linkage
The quantitative understanding of the linkage is low. This is a consequence of it not being possible to measure the upstream and the downstream events in the same study.
Response-response relationshipThe difficulties in extrapolating potency from in vitro to in vivo studies were exemplified by a comparison of the effects of pyrifluquinazon and bisphenol C in vitro and in utero. In vitro, bisphenol C antagonized the androgen receptor with a much higher potency than pyrifluquinazon, but in vivo the potencies were reversed with pyrifluquinazon exposure leading to NR at lower exposure levels than bisphenol C (Gray et al., 2019).
Time-scaleAR activation operates on a time-scale of minutes. The AR is a ligand-activated nuclear receptor and transcription factor. Upon ligand binding a conformational change and subsequent dimerization of the AR takes place within 3-6 minutes (Schaufele et al., 2005). Nuclear translocation (Nightingale et al., 2003) and promoter interactions occur within 15 minutes of ligand binding, and RNA polymerase II and coactivator recruitment are then proposed to occur transiently with cycles of approximately 90 minutes (Kang et al., 2002).
For the downstream event, the time-scale for observing a measurable effect on nipple/areola retention is closer to days and weeks, depending on species. For instance, in mice the nipple anlage are responsive to androgen action at embryonic day 13-15, while a sexual dimorphism of the nipples/areolas can first be observed after birth (Imperato-McGinley et al., 1986) .
Known modulating factorsA well established modulating factor for androgen action is genetic variations in the AR, which decrease the function of the receptor. For example, longer CAG repeat lengths have been associated with decreased AR activation (Chamberlain et al., 1994; Tut et al., 1997). Rat strain is another important modulating factor, with studies showing that the Long-Evans Hooded rat is less sensitive to nipple/areola retention than the Sprague-Dawley rat (Wolf et al., 1999; You et al., 1998)
Known Feedforward/Feedback loops influencing this KERNot relevant for this KER.
References
Amato, C. M., Yao, H. H.-C., & Zhao, F. (2022). One Tool for Many Jobs: Divergent and Conserved Actions of Androgen Signaling in Male Internal Reproductive Tract and External Genitalia. Frontiers in Endocrinology, 13. https://doi.org/10.3389/fendo.2022.910964
Andersson, S., & Russell, D. W. (1990). Structural and biochemical properties of cloned and expressed human and rat steroid 5 alpha-reductases. Proceedings of the National Academy of Sciences, 87(10), 3640–3644. https://doi.org/10.1073/pnas.87.10.3640
Andrade, A. M., Grande, S. W., Talsness, C. E., Grote, K., & Chahoud, I. (2006). Developmental exposure to high and low doses of di-(2-ethylhexyl) phthalate (DEHP); Effects on male rat offspring. NAUNYN-SCHMIEDEBERGS ARCHIVES OF PHARMACOLOGY, 372, 100–101.
Barlow, N. J., McIntyre, B. S., & Foster, P. M. D. (2004). Male reproductive tract lesions at 6, 12, and 18 months of age following in utero exposure to di(n-butyl) phthalate. TOXICOLOGIC PATHOLOGY, 32(1), 79–90. https://doi.org/10.1080/01926230490265894
Borch, J., Axelstad, M., Vinggaard, A. M., & Dalgaard, M. (2006). Diisobutyl phthalate has comparable anti-androgenic effects to di-n-butyl phthalate in fetal rat testis. Toxicology Letters, 163(3), 183–190. https://doi.org/10.1016/j.toxlet.2005.10.020
Borch J, Ladefoged O, Hass U, & Vinggaard AM. (2004). Steroidogenesis in fetal male rats is reduced by DEHP and DINP, but endocrine effects of DEHP are not modulated by DEHA in fetal, prepubertal and adult male rats. Reproductive Toxicology (Elmsford, N.Y.), 18(1), 53–61. https://doi.org/10.1016/j.reprotox.2003.10.011
Chamberlain, N. L., Driver, E. D., & Miesfeld, R. L. (1994). The length and location of CAG trinucleotide repeats in the androgen receptor N-terminal domain affect transactivation function. Nucleic Acids Research, 22(15), 3181–3186. https://doi.org/10.1093/nar/22.15.3181
Christiansen S, Scholze M, Dalgaard M, Vinggaard AM, Axelstad M, Kortenkamp A, & Hass U. (2009). Synergistic disruption of external male sex organ development by a mixture of four antiandrogens. Environmental Health Perspectives, 117(12), 1839–1846. https://doi.org/10.1289/ehp.0900689
Christiansen S, Boberg J, Axelstad M, Dalgaard M, Vinggaard AM, Metzdorff SB, & Hass U. (2010). Low-dose perinatal exposure to di(2-ethylhexyl) phthalate induces anti-androgenic effects in male rats. Reproductive Toxicology (Elmsford, N.Y.), 30(2), 313–321. https://doi.org/10.1016/j.reprotox.2010.04.005
Clewell, R. A., Thomas, A., Willson, G., Creasy, D. M., & Andersen, M. E. (2013). A dose response study to assess effects after dietary administration of diisononyl phthalate (DINP) in gestation and lactation on male rat sexual development. REPRODUCTIVE TOXICOLOGY, 35, 70–80. https://doi.org/10.1016/j.reprotox.2012.07.008
Culty, M., Thuillier, R., Li, W., Wang, Y., Martinez-Arguelles, D. B., Benjamin, C. G., Triantafilou, K. M., Zirkin, B. R., & Papadopoulos, V. (2008). In Utero Exposure to Di-(2-ethylhexyl) Phthalate Exerts Both Short-Term and Long-Lasting Suppressive Effects on Testosterone Production in the Rat1. Biology of Reproduction, 78(6), 1018–1028. https://doi.org/10.1095/biolreprod.107.065649
Euling, S. Y. (2002). Response-Surface Modeling of the Effect of 5alpha-Dihydrotestosterone and Androgen Receptor Levels on the Response to the Androgen Antagonist Vinclozolin. Toxicological Sciences, 69(2), 332–343. https://doi.org/10.1093/toxsci/69.2.332
Fussell KC, Schneider S, Buesen R, Groeters S, Strauss V, Melching-Kollmuss S, & van Ravenzwaay B. (2015). Investigations of putative reproductive toxicity of low-dose exposures to flutamide in Wistar rats. Archives of Toxicology, 89(12), 2385–2402. https://doi.org/10.1007/s00204-015-1622-6
Goldman AS, Shapiro B, & Neumann F. (1976). Role of testosterone and its metabolites in the differentiation of the mammary gland in rats. Endocrinology, 99(6), 1490–1495. https://doi.org/10.1210/endo-99-6-1490
Goldspiel, B. R., & Kohler, D. R. (1990). Flutamide: An Antiandrogen for Advanced Prostate Cancer. DICP, 24(6), 616–623. https://doi.org/10.1177/106002809002400612
Gray LE Jr, Ostby J, Furr J, Price M, Veeramachaneni DN, & Parks L. (2000). Perinatal exposure to the phthalates DEHP, BBP, and DINP, but not DEP, DMP, or DOTP, alters sexual differentiation of the male rat. Toxicological Sciences : An Official Journal of the Society of Toxicology, 58(2), 350–365. https://doi.org/10.1093/toxsci/58.2.350
Gray LE Jr, Barlow NJ, Howdeshell KL, Ostby JS, Furr JR, & Gray CL. (2009). Transgenerational effects of Di (2-ethylhexyl) phthalate in the male CRL:CD(SD) rat: added value of assessing multiple offspring per litter. Toxicological Sciences : An Official Journal of the Society of Toxicology, 110(2), 411–425. https://doi.org/10.1093/toxsci/kfp109
Gray, L. E., Furr, J. R., Conley, J. M., Lambright, C. S., Evans, N., Cardon, M. C., Wilson, V. S., Foster, P. M., & Hartig, P. C. (2019). A Conflicted Tale of Two Novel AR Antagonists in Vitro and in Vivo: Pyrifluquinazon Versus Bisphenol C. Toxicological Sciences, 168(2), 632–643. https://doi.org/10.1093/toxsci/kfz010
Greene, R. R., Burrill, M. W., & Ivy, A. C. (1941). Experimental intersexuality: The effects of combined estrogens and androgens on the embryonic sexual development of the rat. Journal of Experimental Zoology, 87(2), 211–232. https://doi.org/10.1002/jez.1400870203
Hannas, B. R., Furr, J., Lambright, C. S., Wilson, V. S., Foster, P. M. D., & Gray, L. E. (2011). Dipentyl Phthalate Dosing during Sexual Differentiation Disrupts Fetal Testis Function and Postnatal Development of the Male Sprague-Dawley Rat with Greater Relative Potency than Other Phthalates. Toxicological Sciences, 120(1), 184–193. https://doi.org/10.1093/toxsci/kfq386
Hannas, B. R., Lambright, C. S., Furr, J., Evans, N., Foster, P. M. D., Gray, E. L., & Wilson, V. S. (2012). Genomic Biomarkers of Phthalate-Induced Male Reproductive Developmental Toxicity: A Targeted RT-PCR Array Approach for Defining Relative Potency. Toxicological Sciences, 125(2), 544–557. https://doi.org/10.1093/toxsci/kfr315
Hass U, Scholze M, Christiansen S, Dalgaard M, Vinggaard AM, Axelstad M, Metzdorff SB, & Kortenkamp A. (2007). Combined exposure to anti-androgens exacerbates disruption of sexual differentiation in the rat. Environmental Health Perspectives, 115, 122–128. https://doi.org/10.1289/ehp.9360
Hass U, Boberg J, Christiansen S, Jacobsen PR, Vinggaard AM, Taxvig C, Poulsen ME, Herrmann SS, Jensen BH, Petersen A, Clemmensen LH, & Axelstad M. (2012). Adverse effects on sexual development in rat offspring after low dose exposure to a mixture of endocrine disrupting pesticides. Reproductive Toxicology (Elmsford, N.Y.), 34(2), 261–274. https://doi.org/10.1016/j.reprotox.2012.05.090
Hellwig J, van Ravenzwaay B, Mayer M, & Gembardt C. (2000). Pre- and postnatal oral toxicity of vinclozolin in Wistar and Long-Evans rats. Regulatory Toxicology and Pharmacology : RTP, 32(1), 42–50. https://doi.org/10.1006/rtph.2000.1400
Holmer, M. L., Zilliacus, J., Draskau, M. K., Hlisníková, H., Beronius, A., & Svingen, T. (2024). Methodology for developing data-rich Key Event Relationships for Adverse Outcome Pathways exemplified by linking decreased androgen receptor activity with decreased anogenital distance. Reproductive Toxicology, 128, 108662. https://doi.org/10.1016/j.reprotox.2024.108662
Hosokawa S, Murakami M, Ineyama M, Yamada T, Yoshitake A, Yamada H, Miyamoto J. The affinity of procymidone to androgen receptor in rats and mice. J Toxicol Sci. 1993 May;18(2):83-93. doi: 10.2131/jts.18.83. PMID: 8331696.
Howdeshell, K. L., Furr, J., Lambright, C. R., Rider, C. V., Wilson, V. S., & Gray, L. E. (2007). Cumulative Effects of Dibutyl Phthalate and Diethylhexyl Phthalate on Male Rat Reproductive Tract Development: Altered Fetal Steroid Hormones and Genes. Toxicological Sciences, 99(1), 190–202. https://doi.org/10.1093/toxsci/kfm069
Imperato-McGinley J, Binienda Z, Gedney J, & Vaughan ED Jr. (1986). Nipple differentiation in fetal male rats treated with an inhibitor of the enzyme 5 alpha-reductase: definition of a selective role for dihydrotestosterone. Endocrinology, 118(1), 132–137. https://doi.org/10.1210/endo-118-1-132
Jarfelt K, Dalgaard M, Hass U, Borch J, Jacobsen H, & Ladefoged O. (2005). Antiandrogenic effects in male rats perinatally exposed to a mixture of di(2-ethylhexyl) phthalate and di(2-ethylhexyl) adipate. Reproductive Toxicology (Elmsford, N.Y.), 19(4), 505–515. https://doi.org/10.1016/j.reprotox.2004.11.005
Kang, H.-Y., Huang, K.-E., Chang, S. Y., Ma, W.-L., Lin, W.-J., & Chang, C. (2002). Differential Modulation of Androgen Receptor-mediated Transactivation by Smad3 and Tumor Suppressor Smad4. Journal of Biological Chemistry, 277(46), 43749–43756. https://doi.org/10.1074/jbc.M205603200
Kelce, W. R., & Wilson, E. M. (1997). Environmental antiandrogens: Developmental effects, molecular mechanisms, and clinical implications. JOURNAL OF MOLECULAR MEDICINE-JMM, 75(3), 198–207. https://doi.org/10.1007/s001090050104
Klinefelter, G. R., Laskey, J. W., Winnik, W. M., Suarez, J. D., Roberts, N. L., Strader, L. F., Riffle, B. W., & Veeramachaneni, D. N. R. (2012). Novel molecular targets associated with testicular dysgenesis induced by gestational exposure to diethylhexyl phthalate in the rat: a role for estradiol. REPRODUCTION, 144(6), 747–761. https://doi.org/10.1530/REP-12-0266
Kojima, H., Katsura, E., Takeuchi, S., Niiyama, K., & Kobayashi, K. (2004). Screening for estrogen and androgen receptor activities in 200 pesticides by in vitro reporter gene assays using Chinese hamster ovary cells. Environmental Health Perspectives, 112(5), 524–531. https://doi.org/10.1289/ehp.6649
Kratochwil, K. (1977). Development and loss of androgen responsiveness in the embryonic rudiment of the mouse mammary gland. Developmental Biology, 61(2), 358–365. https://doi.org/10.1016/0012-1606(77)90305-0
Kratochwil, K., & Schwartz, P. (1976). Tissue interaction in androgen response of embryonic mammary rudiment of mouse: identification of target tissue for testosterone. Proceedings of the National Academy of Sciences, 73(11), 4041–4044. https://doi.org/10.1073/pnas.73.11.4041
Labrie, F. (1993). Mechanism of action and pure antiandrogenic properties of flutamide. Cancer, 72(12 Suppl), 3816–3827. https://doi.org/10.1002/1097-0142(19931215)72:12+<3816::aid-cncr2820721711>3.0.co;2-3
Martino-Andrade AJ, Morais RN, Botelho GG, Muller G, Grande SW, Carpentieri GB, Leão GM, & Dalsenter PR. (2009). Coadministration of active phthalates results in disruption of foetal testicular function in rats. International Journal of Andrology, 32(6), 704–712. https://doi.org/10.1111/j.1365-2605.2008.00939.x
Matsuura I, Saitoh T, Tani E, Wako Y, Iwata H, Toyota N, Ishizuka Y, Namiki M, Hoshino N, Tsuchitani M, Ikeda Y. Evaluation of a two-generation reproduction toxicity study adding endpoints to detect endocrine disrupting activity using lindane. J Toxicol Sci. (2005) Dec;30 Spec No.:135-161. doi: 10.2131/jts.30.s135. PMID: 16641539.
Mayer, J. A., Foley, J., De La Cruz, D., Chuong, C. M., & Widelitz, R. (2008). Conversion of the nipple to hair-bearing epithelia by lowering bone morphogenetic protein pathway activity at the dermal-epidermal interface. The American Journal of Pathology, 173(5), 1339–1348. https://doi.org/10.2353/AJPATH.2008.070920
McIntyre BS, Barlow NJ, & Foster PM. (2001). Androgen-mediated development in male rat offspring exposed to flutamide in utero: permanence and correlation of early postnatal changes in anogenital distance and nipple retention with malformations in androgen-dependent tissues. Toxicological Sciences : An Official Journal of the Society of Toxicology, 62(2), 236–249. https://doi.org/10.1093/toxsci/62.2.236
Molina-Molina JM, Hillenweck A, Jouanin I, Zalko D, Cravedi JP, Fernández MF, Pillon A, Nicolas JC, Olea N, Balaguer P. Steroid receptor profiling of vinclozolin and its primary metabolites. Toxicol Appl Pharmacol. 2006 Oct 1;216(1):44-54. doi: 10.1016/j.taap.2006.04.005. Epub 2006 Jun 5. PMID: 16750840.
Moore RW, Rudy TA, Lin TM, Ko K, & Peterson RE. (2001). Abnormalities of sexual development in male rats with in utero and lactational exposure to the antiandrogenic plasticizer Di(2-ethylhexyl) phthalate. Environmental Health Perspectives, 109(3), 229–237. https://doi.org/10.1289/ehp.01109229
Mylchreest E, Wallace DG, Cattley RC, & Foster PM. (2000). Dose-dependent alterations in androgen-regulated male reproductive development in rats exposed to Di(n-butyl) phthalate during late gestation. Toxicological Sciences : An Official Journal of the Society of Toxicology, 55(1), 143–151. https://doi.org/10.1093/toxsci/55.1.143
Nightingale, J., Chaudhary, K. S., Abel, P. D., Stubbs, A. P., Romanska, H. M., Mitchell, S. E., Stamp, G. W. H., & Lalani, E.-N. (2003). Ligand Activation of the Androgen Receptor Downregulates E-Cadherin-Mediated Cell Adhesion and Promotes Apoptosis of Prostatic Cancer Cells. Neoplasia, 5(4), 347–361. https://doi.org/10.1016/S1476-5586(03)80028-3
OECD (2013), Guidance Document Supporting OECD Test Guideline 443 on the Extended One-Generational Reproductive Toxicity Test, OECD Series on Testing and Assessment, No. 151, OECD Publishing, Paris, ENV/JM/MONO(2013)10
OECD (2023). Test No. 458: Stably Transfected Human Androgen Receptor Transcriptional Activation Assay for Detection of Androgenic Agonist and Antagonist Activity of Chemicals. OECD Guidelines for the Testing of Chemicals, Section 4, OECD Publishing, Paris, https://doi.org/10.1787/9789264264366-en
OECD (2025a), Test No. 443: Extended One-Generation Reproductive Toxicity Study, OECD Guidelines for the Testing of Chemicals, Section 4, OECD Publishing, Paris, https://doi.org/10.1787/9789264185371-en.
OECD (2025b), Test No. 421: Reproduction/Developmental Toxicity Screening Test, OECD Guidelines for the Testing of Chemicals, Section 4, OECD Publishing, Paris, https://doi.org/10.1787/9789264264380-en.
OECD (2025c), Test No. 422: Combined Repeated Dose Toxicity Study with the Reproduction/Developmental Toxicity Screening Test, OECD Guidelines for the Testing of Chemicals, Section 4, OECD Publishing, Paris, https://doi.org/10.1787/9789264264403-en.
Ogino, Y., Ansai, S., Watanabe, E. et al. Evolutionary differentiation of androgen receptor is responsible for sexual characteristic development in a teleost fish. Nat Commun 14, 1428 (2023). https://doi.org/10.1038/s41467-023-37026-6
Orton, F., Rosivatz, E., Scholze, M., & Kortenkamp, A. (2011). Widely Used Pesticides with Previously Unknown Endocrine Activity Revealed as in Vitro Antiandrogens. ENVIRONMENTAL HEALTH PERSPECTIVES, 119(6), 794–800. https://doi.org/10.1289/ehp.1002895
Ostby J, Monosson E, Kelce WR, & Gray LE Jr. (1999). Environmental antiandrogens: low doses of the fungicide vinclozolin alter sexual differentiation of the male rat. Toxicology and Industrial Health, 15(1), 48–64. https://doi.org/10.1177/074823379901500106
Parks, L. G. (2000). The Plasticizer Diethylhexyl Phthalate Induces Malformations by Decreasing Fetal Testosterone Synthesis during Sexual Differentiation in the Male Rat. Toxicological Sciences, 58(2), 339–349. https://doi.org/10.1093/toxsci/58.2.339
Pedersen, E. B., Christiansen, S., & Svingen, T. (2022). AOP key event relationship report: Linking androgen receptor antagonism with nipple retention. In Current Research in Toxicology (Vol. 3). Elsevier B.V. https://doi.org/10.1016/j.crtox.2022.100085
Saillenfait AM, Sabaté JP, & Gallissot F. (2008). Diisobutyl phthalate impairs the androgen-dependent reproductive development of the male rat. Reproductive Toxicology (Elmsford, N.Y.), 26(2), 107–115. https://doi.org/10.1016/j.reprotox.2008.07.006
Saillenfait AM, Sabaté JP, & Gallissot F. (2009). Effects of in utero exposure to di-n-hexyl phthalate on the reproductive development of the male rat. Reproductive Toxicology (Elmsford, N.Y.), 28(4), 468–476. https://doi.org/10.1016/j.reprotox.2009.06.013
Schaufele, F., Carbonell, X., Guerbadot, M., Borngraeber, S., Chapman, M. S., Ma, A. A. K., Miner, J. N., & Diamond, M. I. (2005). The structural basis of androgen receptor activation: Intramolecular and intermolecular amino–carboxy interactions. Proceedings of the National Academy of Sciences, 102(28), 9802–9807. https://doi.org/10.1073/pnas.0408819102
Schneider S, Kaufmann W, Strauss V, & van Ravenzwaay B. (2011). Vinclozolin: a feasibility and sensitivity study of the ILSI-HESI F1-extended one-generation rat reproduction protocol. Regulatory Toxicology and Pharmacology : RTP, 59(1), 91–100. https://doi.org/10.1016/j.yrtph.2010.09.010
Scholze, M., Taxvig, C., Kortenkamp, A., Boberg, J., Christiansen, S., Svingen, T., Lauschke, K., Frandsen, H., Ermler, S., Hermann, S. S., Pedersen, M., Lykkeberg, A. K., Axelstad, M., & Vinggaard, A. M. (2020). Quantitative in Vitro to in Vivo Extrapolation (QIVIVE) for Predicting Reduced Anogenital Distance Produced by Anti-Androgenic Pesticides in a Rodent Model for Male Reproductive Disorders. Environmental Health Perspectives, 128(11), 117005. https://doi.org/10.1289/EHP6774
Schwartz, C. L., Christiansen, S., Hass, U., Ramhøj, L., Axelstad, M., Löbl, N. M., & Svingen, T. (2021). On the Use and Interpretation of Areola/Nipple Retention as a Biomarker for Anti-androgenic Effects in Rat Toxicity Studies. In Frontiers in Toxicology (Vol. 3). Frontiers Media S.A. https://doi.org/10.3389/ftox.2021.730752
Shimamura M, Kodaira K, Kenichi H, Ishimoto Y, Tamura H, Iguchi T. Comparison of antiandrogenic activities of vinclozolin and D,L-camphorquinone in androgen receptor gene transcription assay in vitro and mouse in utero exposure assay in vivo. Toxicology. 2002 May 24;174(2):97-107. doi: 10.1016/s0300-483x(02)00044-6. PMID: 11985887.
Simard, J., Luthy, I., Guay, J., Bélanger, A., & Labrie, F. (1986). Characteristics of interaction of the antiandrogen flutamide with the androgen receptor in various target tissues. Molecular and Cellular Endocrinology, 44(3), 261–270. https://doi.org/10.1016/0303-7207(86)90132-2
Stoner, E. (1990). The clinical development of a 5α-reductase inhibitor, finasteride. The Journal of Steroid Biochemistry and Molecular Biology, 37(3), 375–378. https://doi.org/10.1016/0960-0760(90)90487-6
Test No. 458: Stably Transfected Human Androgen Receptor Transcriptional Activation Assay for Detection of Androgenic Agonist and Antagonist Activity of Chemicals. (2023). OECD. https://doi.org/10.1787/9789264264366-en
Tut, T. G., Ghadessy, F. J., Trifiro, M. A., Pinsky, L., & Yong, E. L. (1997). Long Polyglutamine Tracts in the Androgen Receptor Are Associated with Reduced Trans -Activation, Impaired Sperm Production, and Male Infertility 1. The Journal of Clinical Endocrinology & Metabolism, 82(11), 3777–3782. https://doi.org/10.1210/jcem.82.11.4385
Vo TT, Jung EM, Dang VH, Jung K, Baek J, Choi KC, Jeung EB. Differential effects of flutamide and di-(2-ethylhexyl) phthalate on male reproductive organs in a rat model. J Reprod Dev. 2009 Aug;55(4):400-11. doi: 10.1262/jrd.20220. Epub 2009 Apr 13. PMID: 19367084.
Welsh, M., Saunders, P. T. K., Fisken, M., Scott, H. M., Hutchison, G. R., Smith, L. B., & Sharpe, R. M. (2008). Identification in rats of a programming window for reproductive tract masculinization, disruption of which leads to hypospadias and cryptorchidism. JOURNAL OF CLINICAL INVESTIGATION, 118(4), 1479–1490. https://doi.org/10.1172/JCI34241
Welsh, M., Suzuki, H., & Yamada, G. (2014). The Masculinization Programming Window. In O. Hiort & S. F. Ahmed (Eds.), UNDERSTANDING DIFFERENCES AND DISORDERS OF SEX DEVELOPMENT (DSD) (Vol. 27, pp. 17–27). https://doi.org/10.1159/000363609
Wilson, V. S., Lambright, C., Furr, J., Ostby, J., Wood, C., Held, G., & Gray, L. E. (2004). Phthalate ester-induced gubernacular lesions are associated with reduced insl3 gene expression in the fetal rat testis. Toxicology Letters, 146(3), 207–215. https://doi.org/10.1016/j.toxlet.2003.09.012
Wilson, V. S., Howdeshell, K. L., Lambright, C. S., Furr, J., & Earl Gray, L. (2007). Differential expression of the phthalate syndrome in male SpragueDawley and Wistar rats after in utero DEHP exposure. Toxicology Letters, 170(3), 177–184. https://doi.org/10.1016/j.toxlet.2007.03.004
Wolf C Jr, Lambright C, Mann P, Price M, Cooper RL, Ostby J, Gray LE Jr. Administration of potentially antiandrogenic pesticides (procymidone, linuron, iprodione, chlozolinate, p,p'-DDE, and ketoconazole) and toxic substances (dibutyl- and diethylhexyl phthalate, PCB 169, and ethane dimethane sulphonate) during sexual differentiation produces diverse profiles of reproductive malformations in the male rat. Toxicol Ind Health. 1999 Jan-Mar;15(1-2):94-118. doi: 10.1177/074823379901500109. PMID: 10188194.
Wolf CJ, LeBlanc GA, Ostby JS, & Gray LE Jr. (2000). Characterization of the period of sensitivity of fetal male sexual development to vinclozolin. Toxicological Sciences : An Official Journal of the Society of Toxicology, 55(1), 152–161. https://doi.org/10.1093/toxsci/55.1.152
Wolf CJ, LeBlanc GA, & Gray LE Jr. (2004). Interactive effects of vinclozolin and testosterone propionate on pregnancy and sexual differentiation of the male and female SD rat. Toxicological Sciences : An Official Journal of the Society of Toxicology, 78(1), 135–143. https://pubmed.ncbi.nlm.nih.gov/14736997/
Wong, C. I., Kelce, W. R., Sar, M., & Wilson, E. M. (1995). Androgen Receptor Antagonist versus Agonist Activities on the Fungicide Vinclozolin Relative to Hydroxyflutamide. Nucleic Acids, Protein Synthesis, and Molecular Genetics, 270(34), 19998–20003.
Wood, A. J. J., & Rittmaster, R. S. (1994). Finasteride. New England Journal of Medicine, 330(2), 120–125. https://doi.org/10.1056/NEJM199401133300208
You, L., Casanova, M., Archibeque-Engle, S., Sar, M., Fan, L.-Q., & Heck, A. (1998). Impaired Male Sexual Development in Perinatal Sprague-Dawley and Long-Evans Hooded Rats Exposed in Utero and Lactationally to p,p’-DDE. In TOX1COLOGICAL SCIENCES (Vol. 45). https://academic.oup.com/toxsci/article/45/2/162/1653877